
Pancreatic beta-cell Protective effect of Chrysophyllum albidum leave extract in Streptozotocin- induced diabetic Rats
- Chika Anna Idaguko
- Agnes Akudo Nwakanma
- 168-182
- Jan 14, 2024
- Physical Education
Pancreatic Beta-Cell Protective Effect of Chrysophyllum Albidum Leave Extract in Streptozotocin- Induced Diabetic Rats
*Chika Anna Idaguko1, Agnes Akudo Nwakanma2
1Department of Anatomy, Faculty of Basic Medical Sciences, Edo State University. Uzairue. Edo State. Nigeria
2Department of Anatomy, Faculty of Basic Medical Sciences, Chukwuemeka Odumegwu Ojukwu University, Uli, Nigeria
*Corresponding Author
DOI: https://doi.org/10.51584/IJRIAS.2023.81216
Received: 24 November 2023; Revised: 08 December 2023; Accepted: 13 December 2023; Published: 13 January 2024
ABSTRACT
Introduction: Chrysophyllum albidum (C. albidum) is a plant that is reported to have antimicrobial, antioxidant, anti-inflammatory and wound healing activities.
Aim: This work was to show that Chrysophyllum albidum extracts protect pancreatic beta-cells from streptozotocin-induced diabetes in Sprague-Dawley rats.
Method: A total of thirty Sprague Dawley rats were used. Group I served as the control with 5 rats while diabetes was induced by injecting a single dose of STZ (50 mg/kg BW) into 25 rats of 5 rats per group. After 3 days of induction of diabetes, the diabetic animals were treated for 28 days with ethanoic extract of C. albidum leave as groups IV, V and VI (125, 250, and 500 mg/kg) respectively, group III was given glibenclamide (2 mg/kg) orally. The blood glucose levels were monitored at regular intervals during the experiment. At the end of the study, the rats were sacrificed and the blood samples were collected for biochemical analysis; and the histology of the pancreatic tissue was conducted.
Results: C. albidum improved, in a dose-dependent manner, the survival and function of rat pancreatic beta-cells from STZ-induced diabetic mediated loss of cell viability and impairment of insulin secretion. Additionally, C. albidum demonstrated potential in reducing TG, LDL-C, TC, and increasing HDL-C. The extract increased the activities of CAT, SOD, and GSH. Finally, C. albidum improved the pancreatic cell injury better than glibenclamide, a medication that is routinely prescribed to treat diabetes. The histological analysis of the pancreas revealed an increase in the number, size, and regeneration of β-cell of islets of Langerhans.
Conclusion: These findings suggested that C. albidum induces insulin secretion that protects beta-cells from oxidative stress-related diabetes.
Keywords: Diabetes, Hyperglycemia, Chrysophyllum albidum, Streptozotocin, glibenclamide.
INTRODUCTION
Diabetes is a multifactorial illness defined by insulin resistance, hyperglycemia, or a combination of these factors [1]. Diabetic dyslipidemia, is another name for the pattern of high triglycerides and low-density lipoprotein (LDL) that is commonly seen in people with diabetes [2] Diabetes-related dyslipidemia is one way that the metabolic syndrome might contribute to the early onset of cardiovascular disease. Hence, one of the leading causes of morbidity and death, is cardiovascular disease; and is rising annually, especially high in people with Type 2 diabetes mellitus [3]. Blood lipid levels are affected by diabetes due to the connection between lipid metabolism and glucose metabolism. Lipoprotein abnormalities are frequently seen in type 2 diabetes [4]. Diabetes frequently results in insulin resistance, which can raise the secretion of very-low-density lipoproteins (VLDLs). Therefore, poor glycemic management raises serum levels of VLDL, TG, and low-density lipoprotein (HDL) cholesterol in people with Type 1 and Type 2 diabetes mellitus. Dyslipidemia is thought to affect between 30 and 60 percent of people with Type 2 diabetes [5]. T1D is caused by a mix of genetic and environmental factors that trigger autoimmune responses that result in the build-up of macrophages and lymphocytes in the islets and the release of pro-inflammatory cytokines such TNF-α, IL-1β, and IFN-α. These cytokines, together with excessive production of reactive oxygen species (ROS) and reactive nitrogen species (RNS) trigger intracellular signaling pathways that encourage autophagy, apoptosis, or necroptosis [6]. In people with type 2 diabetes, increased inflammatory cytokines, hyperglycemia, and hyperlipidemia are caused by insulin resistance and decreased glucose-induced insulin production. In T2D, insulin resistance and impaired glucose-induced insulin secretion lead to sustained hyperglycemia, hyperlipidemia, and elevated inflammatory cytokines These cause lysosomal instability, oxidative stress, and stress on the endoplasmic reticulum (ER), which in turn cause cell death by necroptosis, autophagy, or apoptosis [7]. Therefore, a fundamental factor in both forms of diabetes is beta-cell dysfunction brought on by hyperglycemia-induced reactive oxygen species (ROS) formation [8]. Genetic variables like c-Kit and PPARγ work in concert with a variety of environmental circumstances, such as reduced energy expenditure, increased caloric intake, and excessive ingestion of saturated fat, to create pathological abnormalities that lead to beta cell dysfunction [9]. Insulin resistance, inflammation, immunological dysregulation, elevated sympathetic tone, decreased incretin level/effect, and oxidative stress are a few of these changes [10].
In underdeveloped nations, medicinal plants are sometimes the sole accessible source of economical healthcare and are used extensively in the creation of preventative, controlling, and curative medications for humans [11]. In contrast to oral hypoglycemic medications, which are more likely to cause frequent adverse side effects like mild hypoglycemia and gastrointestinal issues, medicinal plants have attracted a lot of attention recently for the treatment of diabetes mellitus [12]. However, as compared to the oral hypoglycemic medication, medicinal plants are less expensive, more effective, and have fewer adverse effects [13]. Marles and Farnsworth had estimated that approximately 1200 medicinal plant are been used to treat diabetes traditionally [14]. Hence, traditional medicine practices and as evident by ethnopharmacological knowledge have gained attention as potential antidiabetic therapeutics to restore β-cell functions and mass due to their multifaceted ability as in targeting different aspects of the pathways in β-cell regeneration and function [15]. Many mechanisms underlie the antidiabetic action of medicinal plants and phytochemicals, such as decreased intestinal absorption of glucose, inhibition of hepatic glucose synthesis, increased tissue uptake of glucose, renal glucose reabsorption, inhibition of insulin degradative processes, stimulation of islets of Langerhans beta cells for insulin secretion, reduction of insulin resistance, and pancreatic beta cell regeneration or repair through increased islets of Langerhans cell size and number [16,17].
Chrysophyllum albidum know as Africa Star apple, is a species of lowland rain forest tree that grows to a maximum height of 25 to 37 meters at maturity and a girth ranging from 1.5 to 2 meters. It is a member of the Sapotaceae family [18]. The various plant parts of C. albidum are used to treat stomachaches, diarrhea, skin illnesses and as ointments to cure vaginal and dermatological infections. Additionally employed to promote wound healing, stop bleeding from new wounds, and prevent the proliferation of known wound pollutants by microorganisms [19]. The phytochemicals present in C. albidum are a wide range of substances, including flavonoids, glucose, alkaloids, tannins, terpenoids, reducing sugar and cardiac glucosides [20]. These phytochemicals, either individually or combined, may be responsible for the antioxidant properties of C. albidum. Chrysophyllum albidum has been utilized for both medical and nutritional purposes; and it has been linked to a lower incidence of a number of chronic pathological illnesses, such as diabetes, obesity, and cardiovascular diseases [21]. It is commonly known that C. albidum possesses antioxidant qualities [22,23]. However, the literature indicates that there is no specific evidence to support the antidiabetic effect of C. albidum apart from the phytochemicals. The goal of the current study was to determine whether C. albidum’s ethanolic leaf extract might protect the beta cells from STZ-induced diabetes.
MATERIALS AND METHODS
Collection of plant material
Chrysophyllum albidum leaves were collected /from the Monastery located in Elele, River State, Nigeria. Mr. O.O. Oyebanji, a taxonomist at Department of Botany, University of Lagos, Nigeria, identified the leaves. A voucher specimen (LUH 7458) was assigned to the leave and kept in the Botanical Survey of UNILAG’s herbarium. Lagos State. Nigeria
Preparation of test sample
The leaves of Chrysophyllum albidum were finely chopped and left to dry in the shade. Approximately 2 kilograms of the powdered material that had dried was soaked in 5 liters of ethanol in a gas jar for 96 hours at room temperature. Next, the extract was filtered through muslin cloth and Whatman No. 1 filter paper. The ethanol extract was concentrated at 50°C using a rotary evaporator (Rotavapor® R-215, BUCHI Corporation- Switzerland) and further concentrated in an oven at 40°C. This resulted in 168 grams of extract, giving 21.6 percent of the yield. The extract was then stored in a sterilized glass universal bottle and refrigerated at 4°C until used.
Ethical approval
All authors hereby declare that “Principles of laboratory animal care” [24] were followed. All experiments were examined and approved by the College of Medicine University of Lagos Health Research Committee (CMUL/HREC/05/16/011). Animal handling and experiments were performed in strict adherence with the norms of the institutional animal ethics committee.
Animals
Mature male Sprague-Dawley rats weighing 170-200 g were obtained from the National Institute of Medical Research’s (NIMR) animal house. Rats were kept in regular environmental conditions, with a 12:12 light: dark cycle, in polypropylene cages lined with husk. The animals were given at least fourteen days to acclimate before being employed in the experiment. Ad libitum water and a standard pellet diet were provided to the animals.
Chemicals
Glibenclamide and Streptozotocin was purchased from Sigma Chemical Co. (St Louis, Missouri, USA). D-(+)-Glucose Sigma- Aldrich. France (Lot # SZBF1200V). Rat Insulin (INS) ELISA Kit (catalog no: WAR-617) was purchased from Wkea Med Supplies Corp, China. The protocol used for the analysis was as described by the kit producers. All other reagents and chemicals were of analytical grade.
Blood glucose level analysis
Blood Glucose Level (BGL) was measured with Accu-Check Active® (Roche Diagnostic, Mannheim, Germany) active glucose strips and test meter; BGL expressed as mg/dl, using blood obtained from the tail vein of overnight fasted rats.
Evaluation of antidiabetic activity
Induction of diabetes: Streptozotocin (STZ) was administered intraperitoneally (i.p.) to rats at a dose of 50 mg/kg body weight, which was freshly dissolved in 0.1 M cold citrate buffer with a pH of 4.5. Blood samples were taken 72 hours later, and blood glucose levels were measured to confirm the onset of diabetes. In this experiment, rats exhibiting hyperglycemia (blood glucose levels greater than 300 mg/dl) were employed [25]
Treatment model
The rats were divided into six groups of 5 animals (n = 5) each as below:
Group I- Normal control (received distilled water 10 ml/kg b.w.)
Group II- Diabetic control untreated (received distilled water 10 ml/kg b.w.)
Group III- Diabetic treated with standard drug glibenclamide (2 mg/kg/day)
Group IV- Diabetic treated with C. albidum (250 mg/kg/day)
Group V- Diabetic treated with C. albidum (500 mg/kg/day)
Group VI- Diabetic treated with C. albidum (1000 mg/kg/day)
Estimation of plasma glucose
The fasting plasma glucose levels of overnight fasted rats were recorded on 0, 3, 7, 14, 21 and 28 days of the study. Blood glucose was taken from the tail vein of the rats and this was continued for 28 consecutive days using glucose strips, and a glucometer Accu-Check Active® (Roche Diagnostic, Mannheim, Germany). Body weight of all experimental animals was recorded using a digital weighing scale.
Determination of insulin
After 28 days treatment with the extract, and glibenclamide. Before the sacrifice, blood was collected by retro orbital sinus at 0 minute for insulin assay. Glucose load 2 g/kg BW was given to the rats; 30 minutes later blood was collected by retro orbital sinus to determine the insulin assay [23].
Assays of Enzymatic and non-Enzymatic Anti-oxidants
Catalase was estimated as described by [26]. Determination of superoxide dismutase (SOD) as estimated by [27]. The reduced glutathione (GSH) determined by the method of [28]. The malondialdehyde (MDA) level was determined by [29].
Determination of Lipid profile
Serum total cholesterol (TC) was analyzed by enzymatic method (30), Triglyceride (TG) was determined by enzymatic method [31]. High Density Lipoprotein (HDL-c) was analyzed using precipitation method [32], and Low Density Lipoprotein (LDL-c) concentration was determined according to Friedewald et al [33] equation.
Histopathological study
The animals were sacrificed using diethyl ether anesthesia. The pancreas was swiftly cleansed in ice-cold saline and blotted on ash-free filter paper following its dissection. For histological study, a portion of pancreatic tissue was preserved in 10% neutral formalin fixative solution. The steps involved in tissue processing are fixation, dehydration, cleaning, and wax embedding. Block slices were then cut at a thickness of 5 µm, and they were stained with eosin and hemoglobin. The other portion of the pancreas was stained for beta cell stain using Modified aldehyde fuchsin staining technique to highlight beta cells which stained purple-violet [34].
STATISTICAL ANALYSIS
The results were expressed with GraphPad Prism software, version 5.0 (GraphPad Software, La Jolla, CA, USA) and One way analysis of variance (ANOVA) with Bonferroni post-hoc test. The level of significance was set at (P < 0.05). The results were presented as mean ± standard deviation (SD).
RESULTS
Fasting blood glucose level of the ethanolic extract of C. albidum
The results from the study clearly indicate that the ethanolic extract of the leave of C. albidum significantly (P < 0.05) decrease the blood glucose levels from day 14 and this reduction was maintained throughout the 28th days of the treatment, which is comparable to group III. The fasting blood glucose (FBG) of the control (group I) remained unchanged during the course of the study. However, the FBG of the diabetic control (group II) remained elevated throughout the experimental period (Fig. 1)
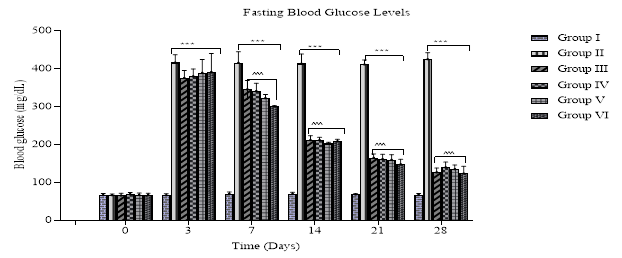
Fig. 1: Bar chart showing the FBG level of rats treated with ethanolic leaf extract of C. albidum. Values represent Mean ± SD (n = 5), *** P < 0.001 compared with control: ^^^ P < 0.001 compared with diabetic control.
Changes of serum insulin
After 28 days of treatment; at 0 minute, the insulin levels of the treated groups were increase accept for the diabetic control II that decreased. 30 minutes after the oral glucose load of 2 mg/kg, there was a significant (P < 0.05) increase in the insulin levels in those administered with 125, 250 and 500 mg/kg of C. albidum, i.e. groups IV, V and VI respectively, when compared to the diabetic control II. (Fig. 2).
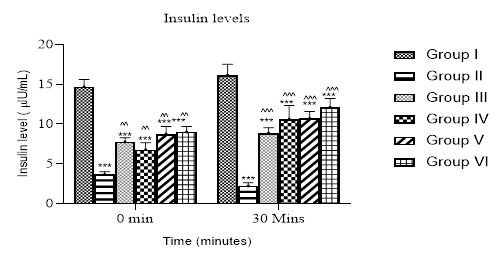
Fig. 2: Bar chart showing insulin level in rats treated with ethanolic leaf extract of C. albidum at 0 minute before glucose load and 30 minutes after glucose load. Values represent Mean ± SD (n = 5), *** P < 0.001 compared with control: ^^ P < 0.01 when compared with diabetic control in 0 minute: ^^^ P < 0.001 when compared with diabetic control in 30 minutes.
At the end of the experiment, there was a statistically significant (P < 0.05) decrease in reduced glutathione (GSH), superoxide dismutase (SOD) and catalase (CAT) activities while malondialdehyde (MDA) content increased in the diabetic group II when compared to the control group I. The GSH activities and MDA content in the groups administered with 125, 250 and 500 mg/kg of C. albidum were comparable to the group III; moreso these groups showed no significant difference when compared to the normal control group I. However, there was significant (P < 0.05) increase of SOD and CAT when compared to the diabetic control group II (Table I).
Table I: Effect Of Ethanolic Leaf Extract of C. Albidum on Oxidative Stress Parameters
| GROUPS | DOSE
mg/kg |
GSH
(U/mg pro) |
SOD
(U/mg pro) |
CAT
(U/mg pro) |
MDA
(U/mg pro) |
| I | 10 ml | 1.63±0.25 | 2.73±0.01 | 22.84±0.11 | 0.20± 0.08 |
| II | 10ml | 0.36±0.11* | 1.50±0.12* | 6.71±0.30* | 1.40±0.00* |
| III | 2 | 1.60±0.21^ | 2.25±0.01*^ | 21.00±0.47*^ | 0.19± 0.00^ |
| IV | 125 | 1.50±0.22^ | 2.81±0.0.1*^ | 11.79±0.80*^ | 0.19±0.05^ |
| V | 250 | 1.63±0.08^ | 3.66±0.09*^ | 18.18±0.49*^ | 0.17±0.05^ |
| VI | 500 | 1.62±0.28^ | 2.55±0.02*^ | 15.83±0.90*^ | 0.20±0.07^ |
Values expressed mean ± SD (n = 5). *(P < 0.05) compared with control. ^ (P < 0.05) compared with diabetic control.
Lipid profile
Diabetic control (group II) showed a significant (P < 0.05) increase in the levels of total cholesterol (TC), triglyceride (TG), low density lipoprotein- cholesterol (LDL-C) and a decreased in high density lipoprotein – cholesterol (HDL-C) levels when compared to control group I. Administration of C. albidum leaf extract showed a significant decrease of TC, TG, LDL-C and increase in HDL-C which is comparable to group III. However, LDL-C was high and HDL-C was also low at 125 mg/kg of C. albidum extract when compared to the control group I as illustrated in (Table II).
Table II: Effects Of Ethanolic Leaf Extract Of C. Albidum On Lipid Profile In S-D Rats
| GROUPS
mg/kg |
DOSE
mmol/L |
TC
mmol/L |
TG
mmol/L |
LDL-C
mmol/L |
HDL-C | |
| I | 10 ml | 0.88±0.01 | 0.65±0.20 | 0.41±0.01 | 0.65±0.01 | |
| II | 10 ml | 2.76±0.12* | 1.42±0.07 * | 1.18± 0.01* | 0.27±0.01* | |
| III | 2 | 0.86±0.21^ | 0.59±0.07^ | 0.47± 0.01^ | 0.68±0.01^ | |
| IV | 125 | 0.75±0.03^ | 0.53±0.04^ | 0.81±0.06*^ | 0.53±0.02*^ | |
| V | 250 | 0.79±0.03^ | 0.53±0.09^ | 0.52±0.02^ | 0.64±0.01^ | |
| VI | 500 | 0.87±0.01^ | 0.54±0.06^ | 0.53±0.03^ | 0.63±0.01^ | |
Values expressed as mean ± SD (n = 5). *(P < 0.05) compared with control, ^ (P < 0.05) compared with diabetic group
Histological sections of the pancreas
Group I exhibits a substantial proportion and typical islet of Langerhans morphology, with the islet buried in acinar cells and encased in a fine capsule. The acinar cells stained strongly and were arranged in lobules with prominent nuclei. Group II shows distortion of the islet organization, necrosis, atrophy of the cells, vacuolation in the islet, and reduction in size and quantity of acinar cells; these findings point to the cytotoxic activity of STZ. Group IV exhibits minimal islet cell mass and partial recovery from the damaged islet cells. On the other hand, groups III, V, and VI displayed an almost normal islet cell architecture that was compact and had very little intercellular space (Fig. 3.)
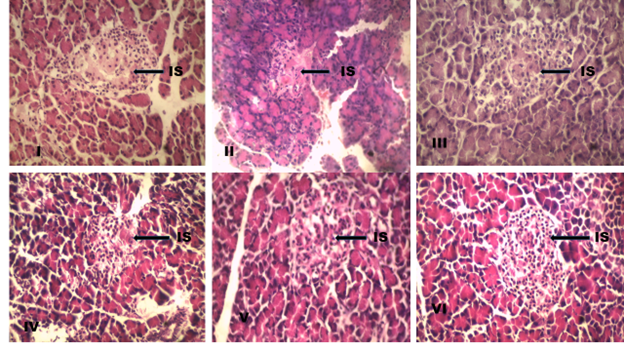
Fig. 3: Photomicrograph of the histological sections of the diabetic study (H and E stain, Mg × 400) IS: (Islet of Langerhans). Group (I) show prominent and well stained islet. Group (II) showing degeneration of islet. Group (IV) show partial recovery of islet. Groups (III, V and VI) shows normal islet organization.
Histological appearance of the beta cells in the study
The photomicrograph showed a compact islet organization, which demonstrated normal morphological features of beta-cells in group 1 (control) (Fig. 4: I A and I B). Group II (diabetic control) showed regular arrangement of beta cells is disturbed, clumping of beta cells, degranulation of the beta cells, beta-cells necrosis and vacuolations (Fig. 4: II A and II B). Group IV (125 mg/kg of C. albidum) highlighted beta-cells (Fig. 4: IV A and IV B). However, group V (250 mg/kg of C. albidum) (Fig. 4: V A and V B) and group VI (500 mg/kg of C. albidum) (Fig. 4: VI A and VI B) showed viable beta-cells at the periphery which is comparable to the group III (glibenclamide) (Fig. 4: III A and III B).
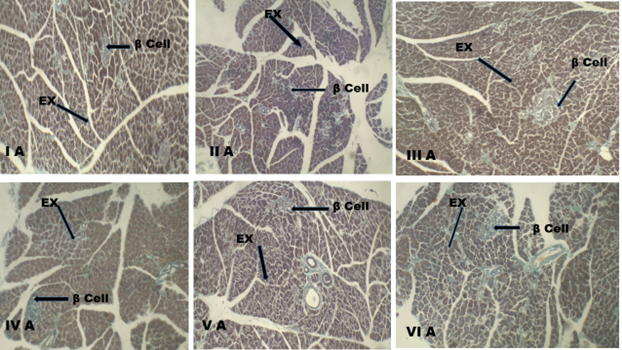
Fig. 4A: Photomicro graph of the pancreatic section, group (I A) normal control showing the exocrine region with beta-cells in the islets of Langerhans. Group (II A) diabetic control showing distorted islet organization. Group (III A) diabetic rat that received glibenclamide (2 mg/kg BW/day) showing beta-cells. Group (IV A) diabetic rat that received 125 mg/kg of C. albidum showing few beta-cells. Group (V A) diabetic rat that received 250 mg/kg of C. albidum showing beta-cells. Group (VI A) diabetic rat that received 500 mg/kg of C. albidum showing beta-cells
EX: Exocrine pancreas, β cell: beta cells stained purple-violet
Stain: Modified Aldehyde Fuchsin, scale bar: 100 µm. Magnification X 100.
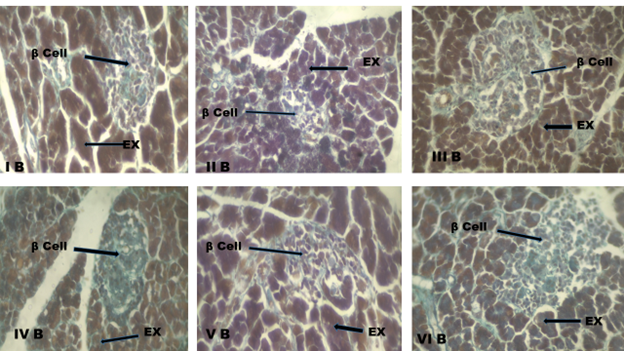
Fig. 4 B: Photomicrograph of the pancreatic section, Group I B: normal control rat showing the exocrine region and beta cells in the islets of Langerhans. Group II B: diabetic control rat showing distorted beta-cells. Group III B: diabetic rat that received glibenclamide (2 mg/kg BW/day) showing beta-cells. Group IV B: diabetic rat that received 125 mg/kg of C. albidum showing beta cells. Group V B: diabetic rat that received 250 mg/kg of C. albidum showing evenly distributed beta-cells. Group VI B: diabetic rat that received 500 mg/kg of C. albidum showing evenly distributed beta-cells.
EX: Exocrine pancreas, β cell: beta cells stained purple-violet.
Stain: Modified Aldehyde Fuchsin, scale bar: 100 µm. Magnification X 400
DISCUSSION
The two primary factors contributing to oxidative stress in diabetes mellitus are hyperglycemia and hyperlipidemia; reactive oxygen species (ROS) produced during this process induce tissue damage and have been proven to have an impact on the two main mechanisms malfunctioning in diabetes: insulin secretion and insulin resistance [35]. As a result, hyperglycemia and hyperlipidemia accompany diabetes. The damage to the pancreatic beta cells may be the cause of the elevated glucose levels in the diabetic control group. Nonetheless, the blood glucose levels were lower in the groups treated with C. albidum. Elevated insulin levels or improved blood glucose transport in peripheral tissue could be the cause of the drop in glucose levels. By increasing muscle glucose absorption and modifying important metabolisms connected to diabetes, C. albidum may also mediate antidiabetic effects [36]. Numerous studies have documented the hyperglycemic characteristics of C. albidum’s various components. According to a study by Ajayi et al. [21], blood glucose in STZ-induced diabetic rats was decreased by C. albidum ethanolic fruit extract at 100 and 200 mg/kg. In STZ-induced diabetic rats, 30 mg/kg of C. albidum’s ethanolic stem bark extract reduced blood glucose levels [37]. On STZ-diabetic Wistar albino rats, the ethanolic fruit-skin extract of C. albidum at 100 and 200 mg/kg decreased the blood glucose level [38]. Streptozotocin and a high-fat diet-induced blood glucose level in diabetics were both considerably (p<0.05) reduced by 5 and 10% of C. albidum pulp powder [39]. Blood glucose was considerably (p < 0.05) decreased by C. albidum ethanol root bark extract [40]. Blood sugar levels are significantly lowered (P<0.05) by C. albidum seed and leaf extracts [41]. In STZ-induced diabetic mice, pretreatment with an ethanolic C. albidum leaf extract at 250 and 500 mg/kg decreased blood glucose levels and complications linked to hyperglycemia [22, 23].
Additionally, blood cholesterol, TG, LDL, and HDL-C levels were all higher in diabetic rats. Since insulin inhibits the hormone-sensitive lipase, the abnormally high quantity of blood lipids in diabetes patients is caused by an increase in the activity of free fatty acids from the peripheral fat depots. Serum fatty acids are produced in excess when a person has diabetes, and the liver converts them into phospholipids and cholesterol [4]. These two compounds may be released into the bloodstream as lipoproteins, together with any extra triglycerides that the liver produces at the same time [4]. Major risk factors for atherosclerosis include hypertriglyceridemia and hypercholesterolemia. Triglycerides are hydrolyzed by lipoprotein lipase when insulin is activated during regular metabolic processes. However, lipoprotein lipase is not activated in an insulin-deficient state, which leads to hypertriglyceridemia [42]. The study showed a reduction in TG, TC, and LDL following C. albidum treatment. The decrease in cholesterol biosynthesis may primarily result from the suppression of the rate-limiting enzyme HMG-CoA reductase and the enhancement of lipolysis through the reduction of hormone-sensitive lipase activity [43]. The administration of plant treatments resulted in a noteworthy increase in HDL-C, suggesting a potential protective effect against cardiovascular disease. The mechanism of this activity maybe through the inhibition of LDL oxidation, facilitation of the reverse cholesterol transport pathway, stimulation of the excretion of accumulated excess cellular cholesterol, and prevention of the production of oxidatively modified LDL particles [44].
One of the primary roles of pancreatic beta cells is the release of insulin, which maintains glucose homeostasis. The primary physiological factor that initiates insulin production is an increase in blood glucose. Once glucose enters beta cells, it raises the ATP to ADP ratio, which closes the KATP channels and depolarizes the cell membrane. In order to trigger the exocytosis of insulin granules, the opens voltage-dependent calcium channels, raises intracellular calcium [Ca2+] leading to the activation of calcium/calmodulin-dependent pathways [45,46]. The suppression of oxidative stress and subsequent inhibition of beta cell damage is one of the primary molecular mechanisms by which plants safeguard the life of beta cells [47]. Beta cells are susceptible to oxidative stress due to their low intracellular antioxidant capacity. Thus, in diabetes, oxidative molecules play a significant role as mediators of beta cell destruction [48]. Additionally, reactive oxygen species are produced as a result of chronic hyperglycemia, which damages β-cells [35].
The antioxidant enzymes produced by the cells themselves, known as endogenous enzymes, serve as a defense against reactive free radicals [49]. The three most important antioxidant enzymes are superoxide dismutase (SOD), catalase (CAT), and GST. O2•–was oxidized by the SOD into hydrogen peroxide (H202), which is then transformed into water (H2O) by the CAT and Fenton reaction. As a result, hazardous species are transformed into innocuous products. GST gets rid of peroxides produced throughout the metabolic process [50]. According to report, enhanced antioxidant ability shields beta cells against autophagy, apoptosis, and necroptosis by preventing ROS generation and lipid peroxidation [51].
When given to diabetic rats, C. albidum boosts the antioxidant capacity of beta cells by lowering MDA and boosting non-enzymatic (reduced glutathione) and enzymatic (superoxide dismutase and catalase) antioxidants. This is consistent with a publication showing antioxidants or free radical scavengers in plant extract can protect beta cells from oxidative damage. The C. albidum extract may have increased serum insulin because it stimulated insulin release or preserved beta cell viability as depicted in Fig. 2. According to a number of studies, flavonoids prevent beta cell dysfunction brought on by cytokines and improve insulin release in response to glucose [52.53]. These advantageous outcomes are mediated by influencing both triggering and amplifying pathways of insulin secretion mechanisms
The study’s histological analysis demonstrated that the extract-treated diabetic rats exhibited variable levels of islet cell recovery; the extract also markedly increased the number of beta cells and restored the pancreatic cells. The regeneration of islet cells, which occurs primarily around 250 and 500 mg/kg of extracts, strongly suggests that stable cells capable of regeneration exist in the islets. Therefore, the ethanolic leaf extract of C. albidum at 250 and 500 mg/kg demonstrated a protective effect that was comparable to the standard medication utilized in this investigation, glibenclamide (2 mg/kg). Thus, β-Cell regeneration can happen through neogenesis from stem cells, or replication of already-existing β-cells and transforming other pancreatic cells into β-cells [15]. Different cell types found in the pancreas, including duct epithelium, acinar cells, centroacinar cells, delta cells, and alpha cells, can give rise to neogenesis [54]. Hormones, growth factors, and other extra-pancreatic activators are necessary for this process to occur [55]. Therefore, the extract might have a regenerative or protective effect on beta cells in the pancreas. The hypoglycemic action of C. albidum may also be explained by the histology findings. Some plants have been reported to improve the function of beta cells in the pancreas [56].
Plant phytoconstituents, including alkaloids, glycosides, phenol, terpenoids, flavonoids, and tannins, have been linked to considerable antioxidant activity and frequently implicated with the antidiabetic properties [57]. However, the flavonoid and phenolic content may possibly be the reason that treatments with ethanolic leaf extracts dramatically improve the degenerative abnormalities of the pancreas. Since flavonoids are intermediate biosynthetic precursors and alpha-amylase inhibitors, they may be able to restore injured beta cells seen in C. albidum. Furthermore, the presence of tannin and polyphenolic chemicals will prevent glucose from being transported by blocking the sodium glucose co-transporter-1 (SGLUT-1) in the rats’ intestines [58].
Polyphenol compounds are suggested to have significant effects on the regulation of oxidative stress through a number of mechanisms [59]. These include reducing inflammation through inactivation of mitogen-activated protein kinase (MAPK) signaling pathways, repairing DNA damages caused by oxidative stress via the nuclear factor erythroid 2-related factor 2 (Nrf2) pathway [60], and decreasing apoptosis through controlling protein kinase B (AKT)/caspase expression [61]. Furthermore, some phytochemicals promote the proliferation and differentiation of progenitor cells involved in protection and regeneration of β-cells [62].
According to [63], Gas Chromatography-Mass Spectrometry (GC-MS) of the C. albidum leaf extract was shown to contain the following compounds, whose activities have been identified; 1,2,3-benzenetriol (Pyrogallol), a polyphenol chemical with anti-inflammatory and antioxidant properties. Triterpene 2(4H)-Benzofuranone 5,6,7,7a-tetrahydro-4,4,7 a-trimethyl having an antidiabetic and antioxidant activities. 4-Acetamidobutyric acid a gamma amino acids with antioxidant properties. 1,2,3-propanetriol, 1-acetate, also known as acetin, having anti-inflammatory, hypoglycemic, hypolipidemic, antidiabetic properties and can also regenerate beta cells. The phenolic compound 4H-Pyran-4-one, 2,3-dihydro-3, 5-dihydroxy-6-methyl having also anti-inflammatory and antioxidant properties. In addition, sucrose a disaccharide having antioxidant, triglycerigenic, atherogenic, and hypercholesterolemic activities. This compounds may be the active components responsible for the protective effect of C. albidum leave extract against beta cell destruction caused by STZ as seen in this study.
Hence, C. albidum increased insulin sensitization by increasing insulin secretion, and beta cell regeneration in STZ- induced diabetic rats. In addition, C. albidum show antihyperglycemic, antioxidant, hypolipidemic properties and has beneficial effects on the histological structure of the islets of Langerhans of the pancreas.
CONCLUSION
According to the results, C. albidum have a positive impact on the function of pancreatic beta cells in STZ-induced animal model, by reducing the blood glucose, antioxidant and antihyperlipidemic activities. Also it increase the secretion of insulin thereby increasing the number and regeneration, of beta cells
ACKNOWLEDGMENTS
The authors which to acknowledge Prof Duru FIO and Prof Oremosu A.A for their assistance during the study.
COMPETING INTERESTS
The authors declares no competing interest
AUTHOR’S CONTRIBUTIONS
Chika. Anna Idaguko designed the protocol, carried out the statistical analysis, and penned the initial draft of the paper. Akudo Agnes Nwakanma oversaw the literature searches and the study’s analyses. The final manuscript was read and approved by both authors.
REFERENCES
- Saeedi, P, Petersohn I, Salpea P, Malanda B, Karuranga S, Unwin N, et al (2019). Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas, 9(th) edition. Diabetes Res Clin Pract. 157:107843.
- Lokpo, S.Y, Laryea R, Osei-Yeboah J, Owiredu W.K.B.A, Ephraim R.K.D, Adejumo E.N, et al (2022). The pattern of dyslipidaemia and factors associated with elevated levels of non-HDL-cholesterol among patients with type 2 diabetes mellitus in the Ho municipality: A cross sectional study. Heliyon. 8(8):e10279. Available from: https://www.sciencedirect.com/science/article/pii/S2405844022015675
- Ma, C.X, Ma X.N, Guan C.H, Li Y.D, Mauricio D, Fu S.B (2022). Cardiovascular disease in type 2 diabetes mellitus: progress toward personalized management. Cardiovasc Diabetol. 21(1):74. Available from: https://doi.org/10.1186/s12933-022-01516-6
- Dilworth, L, Facey A, Omoruyi F (2021). Diabetes Mellitus and Its Metabolic Complications: The Role of Adipose Tissues. Int J Mol Sci. 22(14):7644.
- Thambiah, CS, Lai LC. (2021). Diabetic dyslipidaemia. Pract Lab Med. 26:e00248. Available from: https://www.sciencedirect.com/science/article/pii/S2352551721000482
- Toren, E, Burnette K.S, Banerjee R.R, Hunter C.S (2021). Partners in Crime : Beta-Cells and Autoimmune Responses Complicit in Type 1 Diabetes Pathogenesis.12:1–19.
- Ghorbani, A, Rashidi R, Shafiee-Nick R (2019). Flavonoids for preserving pancreatic beta cell survival and function: A mechanistic review. Biomed Pharmacother. 111:947–57. Available from: https://www.sciencedirect.com/science/article/pii/S0753332218375711
- Mukai, E, Fujimoto S, Inagaki N (2022). Role of Reactive Oxygen Species in Glucose Metabolism Disorder in Diabetic Pancreatic β-Cells. Biomolecules. 12(9).
- Acosta-Montaño, P, García-González V (2018). Effects of Dietary Fatty Acids in Pancreatic Beta Cell Metabolism, Implications in Homeostasis. Nutrients. 10(4).
- Kosmas, C.E, Bousvarou M.D, Kostara C.E (2023), Papakonstantinou EJ. Insulin resistance and cardiovascular disease. J Int Med Res. 51(3):1–49.
- Eshete, M.A, Molla E.L (2021).. Cultural significance of medicinal plants in healing human ailments among Guji semi-pastoralist people, Suro Barguda District, Ethiopia. J Ethnobiol Ethnomed. 17(1):61. Available from: https://doi.org/10.1186/s13002-021-00487-4
- Mohammed, A (2023). Hypoglycemic Potential of African Medicinal Plants in Diabetic and Non-Diabetic Human Subjects: A Review. Clin Complement Med Pharmacol. 3(2):100081. Available from: https://www.sciencedirect.com/science/article/pii/S2772371223000037
- Rahman, M.M, Dhar P.S, Sumaia, Anika F, Ahmed L, Islam M.R, et al (2022). Exploring the plant-derived bioactive substances as antidiabetic agent: An extensive review. Biomed Pharmacother. 152:113217. Available from: https://www.sciencedirect.com/science/article/pii/S0753332222006060
- Marles, RJ, Farnsworth NR (1995). Antidiabetic plants and their active constituents. Phytomedicine. 2(2):137–89.
- Wickramasinghe, A.S.D, Kalansuriya P, Attanayake A.P (2021). Herbal Medicines Targeting the Improved β-Cell Functions and β-Cell Regeneration for the Management of Diabetes Mellitus. Evid Based Complement Alternat Med. 2021:2920530.
- Przeor, M (2022). Some Common Medicinal Plants with Antidiabetic Activity, Known and Available in Europe (A Mini-Review). Pharmaceuticals (Basel). 15(1):65.
- Hosseini , A, Shafee-Nick R, Ghorbani A (2015). Pancreatic beta cell protection/regeneration with phytotherapy. Brazilian J Pharm Sci. 51(1):1–16.
- Adelani, D, Aduradola M, Osunsina O 2020. Effects of watering levels and light intensities on seeding vigour of African Stat Apple (Chrysophyllum albidum G. DON). J Res For Wildl Environ. 12(2):90–9.
- Ushie, OA, Adamu H.M, Abayeh O.J, Chindo I.Y, Lawal U (2014). Phytochemical Screening of ( chrysophyllum albidum ) Leaf Extracts. J Appl Chem. 2(2):40–6.
- Abiol,a Ogunleye F, Fapohunda O, Nwangwu S (2020) A Review on Medicinal Uses and Pharmacological Activities of African Star Apple (Chrysophyllum albidum). Acta Sci Pharmacol. 1(4).
- Ajayi, A.M, Adedapo A.D..A, Badaki VB, Oyagbemi A.A, Adedapo A.A (2021). Chrysophyllum albidum fruit ethanol extract ameliorates hyperglycaemia and elevated blood pressure in streptozotocin-induced diabetic rats through modulation of oxidative stress, NF-κB and PPAR-γ. Biomed Pharmacother.141:111879. Available from: https://www.sciencedirect.com/science/article/pii/S0753332221006612
- Idaguko, C, A, Oremosu A, Duru A, Awopetu P (2018). Protective Effect of Ethanolic Leaf Extract of Chrysophyllum albidum (Sapotaeae G. Don) on Histological Changes in the Pancreas of Streptozotocin-induced Diabetic Sprague Dawley Rats. J Anat Sci. 9(1):1–6.
- Idaguko, C,A, Duru F, Oremosu A. (2017) .Antioxidant, hypolipidemic and Hypoglycemic Effect of Ethanol Leaf Extract of Chrysophyllum albidum on Streptozotocin- Induced Diabetic Rats. J Anat Sci. 8(1):67–73.
- Guide for the Care and Use of Laboratory Animals. 8TH ed. Washington, DC: National Academies Press; 2011. 15 p.
- Okoduwa, SIR, Umar IA, James DB, Inuwa HM. (2017). Anti-Diabetic Potential of Ocimum gratissimum Leaf Fractions in Fortified Diet-Fed Streptozotocin Treated Rat Model of Type-2 Diabetes. Med (Basel, Switzerland). 4(4).
- Choudhary, R, Saroha AE, Swarnkar PL. (2012). Effect of abscisic acid and hydrogen peroxide on antioxidant enzymes in Syzygium cumini plant. J Food Sci Technol. 49(5):649–52.
- Sun, M, Zigman S. (1978). An improved spectrophotometric assay for superoxide dismutase based on epinephrine autoxidation. Anal Biochem. 90(1):81–9.
- Davies, H, Birt D. F, Schnell R. C. (1984) Direct enzymatic assay for reduced and oxidized glutathione. J Pharmacol Methods. 12(3):191–4. Available from: https://www.sciencedirect.com/science/article/pii/0160540284900597
- D’souza, D, Subhas B. G, Shetty S. R, Balan P (2012). Estimation of serum malondialdehyde in potentially malignant disorders and post-antioxidant treated patients: A biochemical study. Contemp Clin Dent. 3(4):448–51.
- Allain, C. C, Poon L. S, Chan C. S, Richmond W, Fu P. C (1974). Enzymatic determination of total serum cholesterol. Clin Chem. 20(4):470–5.
- Ezeonwumelu, E, Omeodu S. (2019). Serum Lipid Profile , Liver Function Indices and Electrolyte Levels in Diabetics and Subjects with Hepatic Impairment in the University of Port Harcourt Teaching Hospital. J Appl Sci Environ Manag. 23(2):261–268.
- Lopes-Virella, M. F, Stone P, Ellis S, Colwell J. A (1977). Cholesterol determination in high-density lipoproteins separated by three different methods. Clin Chem. 23(5):882–4.
- Friedewald, W. T, Levy R. I, Fredrickson D. S. (1972). Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin Chem. 18(6):499–502.
- Lyon, H, Prentø P (1980). Aldehyde Fuchsin staining of pancreatic B cells. Reproducible high-contrast staining of formalin-fixed and paraffin-embedded material. Histochem J. 12(1):97–105.
- Bhatt,i JS, Sehrawat A, Mishra J, Sidhu IS, Navik U, Khullar N, et al. (2022). Oxidative stress in the pathophysiology of type 2 diabetes and related complications: Current therapeutics strategies and future perspectives. Free Radic Biol Med [Internet].184:114–34. Available from: https://www.sciencedirect.com/science/article/pii/S0891584922001228
- Erukainure, O, Salau V, Xiao X, Matsabisa M, Koorbanally N, Islam M (2021). Bioactive compounds of African star apple (Chrysophyllum albidum G. Don) and its modulatory effect on metabolic activities linked to type 2 diabetes in isolated rat psoas muscle. J Food Biochem [Internet].;45(1):e13576. Available from: https://doi.org/10.1111/jfbc.13576
- Akanji, O, Ogunma G, Oshomoh E, Asuelimen O. (2020). Antidiabetic property of Chrysophyllum albidum extract in Streptozotozin- induced diabetic rats. J Basic Appl Zool. 9(4):230–5.
- Asagba, S. O, Kadiri H. E, Ezedom T. (2019). Biochemical changes in diabetic rats treated with ethanolic extract of Chysophyllum albidum fruit-skin. J Basic Appl Zool. 80:42.
- Oyetayo, F, Akomolafe S, Jegede F, Elekofehinti O, Akinjiyan M, Odeniyi I. (2012). Effect of Chrysophyllum albidum fruit pulp powder on antioxidant and proinflammatory genes in non-diabetic and type 2 diabetic rats. J Diabetes Metab Disord [Internet]. 20(2):1663–74. Available from: https://doi.org/10.1007/s40200-021-00921-0
- Onyeka, C. A, Nwakanma A, Bakare A, Okoko I, Ofoego U, Wali C, et al. (2013) Hypoglycemic, antioxidant and hepatoprotective activities of ethanolic root bark extract of chrysophyllum albidum in alloxan-induced diabetic rats. Bangladesh J Med Sci.12(3):298–304.
- Godwill, E, Unaegbu M, Esther A, Gloria O, Kingsley A, Aiyegoro O, et al (2016). Antioxidant and antidiabetic activities of the seed and leaf extracts of Chrysophyllum albidum. Asian Pacific J Trop Dis. 6(8):642–9.
- Petersen, M. C, Shulman G. I (2018). Mechanisms of Insulin Action and Insulin Resistance. Physiol Rev. 98(4):2133–223.
- Attanayake, A. P, Jayatilaka K. A. P. W, Mudduwa L. K. B, Pathirana C. (2018). Biochemical and Histological Evaluation of Three Selected Medicinal Plant Extracts of Sri Lankan Origin on Dyslipidemia and Oxidative Stress in Alloxan Induced Diabetic Rats. Mahmood T, editor. J Bot Available from: https://doi.org/10.1155/2018/4204519
- Rani, A, Marsche G. A (2023). Current Update on the Role of HDL-Based Nanomedicine in Targeting Macrophages in Cardiovascular Disease. Pharmaceutics. 15
- Gelbach , P. E, Zheng D, Fraser S. E, White K. L, Graham N. A, Finley S. D. (2022). Kinetic and data-driven modeling of pancreatic β-cell central carbon metabolism and insulin secretion. PLOS Comput Biol. 18(10):e1010555. Available from: https://doi.org/10.1371/journal.pcbi.1010555
- Mann , E, Sunni M, Bellin M. D. (2020). Secretion of Insulin in Response to Diet and Hormones I . The Dual Nature of the Pancreas III . Insulin Gene Transcription II . Insulin Structure. Pancreapedia Exocrine Pancreas Knowl Base. 1–17. Available from: doi: 10.3998/panc.2020.16
- Dinić , S, Arambašić Jovanović J, Uskoković A, Mihailović M, Grdović N, Tolić A, et al. (2022). Oxidative stress-mediated beta cell death and dysfunction as a target for diabetes management. Front Endocrinol (Lausanne). 13:1006376.
- Eguch, i N, Vaziri N. D, Dafoe D. C, Ichii H (2021). The Role of Oxidative Stress in Pancreatic β Cell Dysfunction in Diabetes. International Journal of Molecular Sciences. 22.
- Jena, A.B, Samal R.R, Bhol N.K, Duttaroy A.K (2023). Cellular Red-Ox system in health and disease: The latest update. Biomed Pharmacother. 162:114606. Available from: https://www.sciencedirect.com/science/article/pii/S0753332223003943
- Chaudhary, P, Janmeda P, Docea AO, Yeskaliyeva B, Abdull Razis A. F, Modu B, et al. Oxidative stress, free radicals and antioxidants: potential crosstalk in the pathophysiology of human diseases. Front Chem [Internet]. 2023;11. Available from: https://www.frontiersin.org/articles/10.3389/fchem.2023.1158198
- Su , LJ, Zhang J.H, Gomez H, Murugan R, Hong X, Xu D, et al (2019). Reactive Oxygen Species-Induced Lipid Peroxidation in Apoptosis, Autophagy, and Oxid Med Cell Longev. 2019:5080843.
- Zhou, M, Konigsberg W.H, Hao C, Pan Y, Sun J, Wang X (2023). Bioactivity and mechanisms of flavonoids in decreasing insulin resistance. J Enzyme Inhib Med Chem 38(1):2199168. Available from: https://doi.org/10.1080/14756366.2023.2199168
- Lee, S, Chyau C.C, Wang C.P, Wang T.H, Chen J.H, Lin H.H (2020). Flavonoids Identification and Pancreatic Beta-Cell Protective Effect of Lotus Seedpod. Antioxidants. 9
- Li, K, Bian J, Xiao Y, Wang D, Han L, He C, et al (2023). Changes in Pancreatic Senescence Mediate Pancreatic Diseases. International Journal of Molecular Sciences. 24
- Paris, M, Tourrel-Cuzin C, Plachot C, Ktorza A (2004). Review: pancreatic beta-cell neogenesis revisited. Exp Diabesity Res. 5(2):111–21.
- Oliaee, D, Niazkar H, Abbasnezhad.A, Ghorbani M, Alavi Shahri P, Saghaee Shahri S, et al (2020). The Effects of Medicinal Plants on Pancreatic Beta Cells in Diabetes : A Systematic Review of Iranians ’ Contributions. Rev Clin Med. 7(1):30–6.
- Tran, N, Pham B, Le L. (2020). Bioactive Compounds in Anti-Diabetic Plants: From Herbal Medicine to Modern Drug Discovery. Biology (Basel). 9(9).
- Brus, M, Frangež R, Gorenjak M, Kotnik P, Knez Ž, Škorjanc D (2021). Effect of Hydrolyzable Tannins on Glucose-Transporter Expression and Their Bioavailability in Pig Small-Intestinal 3D Cell Model. Molecules. 26(2).
- Rudrapal, M, Khairnar S.J, Khan J, Dukhyil A Bin, Ansari M.A, Alomary M.N, et al. (2022). Dietary Polyphenols and Their Role in Oxidative Stress-Induced Human Diseases: Insights Into Protective Effects, Antioxidant Potentials and Mechanism(s) of Action. Front Pharmacol. 13:806470.
- Krawczyk, M, Burzynska-Pedziwiatr I, Wozniak L.A, Bukowiecka-Matusiak M. (2023). Impact of Polyphenols on Inflammatory and Oxidative Stress Factors in Diabetes Mellitus: Nutritional Antioxidants and Their Application in Improving Antidiabetic Therapy. Biomolecules. 13
- Li, S, Tan, Y, Wang N, Cheung F, Hong, M, Feng Y (2018). The Potential and Action Mechanism of Polyphenols in the Treatment of Liver Diseases. Oxid Med Cell Longev. 2018:8394818.
- Kimani, N, Reuter, H, Kotzé, S.H, Muller C.J.F. (2023). Regeneration of Pancreatic Beta Cells by Modulation of Molecular Targets Using Plant-Derived Compounds: Pharmacological Mechanisms and Clinical Potential. Curr Issues Mol Biol. 45(8):6216–45.
- Idaguko, A, Orabueze I. (2023). Trace element levels and bio-active compounds in ethanolic leaf extract of Chrysophyllum albidum characterised using Gas Chromatography – Mass Spectrometry. J Trace Elem Med Biol. 80:127311. Available from: https://www.sciencedirect.com/science/article/pii/S0946672X23001876