
Molecular Identification of Trichophyton Rubrum Onychomycosis at Health Centres in Benue State, Nigeria
- Magdalene N. Tyona
- Grace M. Gberikon
- Terdzungwe T. Sar
- 309-316
- Apr 14, 2024
- Microbiology
Molecular Identification of Trichophyton Rubrum Onychomycosis at Health Centres in Benue State, Nigeria
Magdalene N. Tyona*1, Grace M. Gberikon2 & Terdzungwe T. Sar3
1,2,3Joseph Sarwuan Tarka University, Makurdi, Benue State, Nigeria
*Correspondence Author
DOI: https://doi.org/10.51584/IJRIAS.2024.90329
Received: 14 January 2024; Accepted: 07 February 2024; Published: 14 April 2024
ABSTRACT
The molecular properties of Trichophyton rubrum associated with onychomycosis in adults attending selected tertiary and secondary health facilities in Benue State, Nigeria were investigated. The prevalence of onychomycosis, co-morbidities, onychomycosis related health histories, and infecting strains of Trichophyton rubrum were considered. Due ethical clearance was obtained, and informed consent granted by participants. The research employed purposive random sampling technique. Three hundred and eighty-four (384) adults aged 18 years and above, with visible symptoms of onychomycosis were recruited. Nail samples were collected, disinfected and examined. Sabouraud Dextrose Agar (SDA) was used for fungal cultures. Laboratory procedures included microscopy and morphological characterization of isolates. Toenails were more infected 210 (68.48%) than fingernails 8 (18.18%). The fungal species with the highest frequency of occurrence was T. rubrum 159 (70.36%), while the least was Candida spp. 19 (8. 49%). Allele-Specific Polymerase chain reaction (PCR) revealed new sub-repeat elements in the non-transcribed region of the rDNA spacer of T. rubrum. The study has documented onychomycosis prevalence rates and helped provide insight into the infecting species in Benue State. This will help develop appropriate treatment and intervention guidelines. Further and advanced molecular typing methods for enhanced understanding of infecting fungal strains is recommended.
Key Words: Onychomycosis, T. rubrum, Allele-Specific PCR, Benue State, non-transcribed region
INTRODUCTION
Onychomycosis is a fungal infection of the nails of significant public health concern globally. Its etiological agents include various dermatophytes (de Berker et al., 2007). Infection can significantly impact patients’ emotional, social, and occupational functioning. Patients feel embarrassed in social and workplace situations, unwilling to expose their hands or feet. Several factors predispose individuals to onychomycosis, and include diabetes mellitus, age, nail trauma, poor peripheral circulation, immunosuppression, tightly fitting shoes, long nails, fixing of artificial nails, going barefoot in public places, sharing shoes, and exposing hands or feet to damp environments (Westerberg and Voyack, 2013).
The prevalence of onychomycosis has been on the rise, underscoring the need for a comprehensive understanding of causative agents and associated risk factors (Baki et al., 2018). Trichophyton rubrum, a dermatophyte known for its proclivity to infect keratinized tissues, is a key contributor to the global burden of onychomycosis (Jazdarehee et al., 2022).
Molecular identification techniques have emerged as pivotal tools in the accurate and precise categorization of infecting fungal strains, allowing for deeper understanding of their genetic makeup and epidemiological patterns. Polymerase chain reaction (PCR) and DNA sequencing, among other molecular methods, help researchers distinguish between infecting fungal species and strains, thus facilitating a targeted approach to diagnosis and treatment (Jackson et al., 1999).
Nigeria, like many other tropical regions, presents an environment conducive to the proliferation of dermatophytes due to factors such as high temperatures and humidity (Yenişehirli et al., 2012). Investigating and understanding the local Benue epidemiology of onychomycosis and its causative agents is crucial for tailoring effective preventive measures and treatment strategies in this specific population.
The molecular identification of Trichophyton rubrum onychomycosis in Benue State is justified by its potential to inform targeted interventions, optimize treatment strategies, and contribute valuable knowledge to the broader understanding of fungal infections in diverse populations. The outcomes of this research have the potential to significantly impact public health policies, clinical practices, and the overall well-being of individuals affected by onychomycosis in Benue State and beyond.
Study Area
The study was carried out at Federal Medical Centre, Makurdi, Benue State University Teaching Hospital, Makurdi, Bishop Murray Medical Center, Makurdi, General Hospital, Otukpo and General Hospital, Gboko and spanned 8 months between February to October 2023.
Benue State lies within the lower River Benue trough in the middle belt region of Nigeria on longitude 7o 47’ and 10o 0’ East and Latitude 6o 25’ and 8o 8’ North (Agada and Nirupama, 2015). Benue state lies within the AW climate and experiences two distinct seasons: the wet and dry seasons. The wet season lasts from April to October with annual rainfall between 100-200mm. The dry season begins in November and ends in March. The yearly temperatures fluctuate between 21-37oC (Adejuwon, 2012).
Ethical Considerations
The study was approved by the respective Ethics Committees of the Benue State Ministry of Health and Federal Medical Centre, Makurdi. Furthermore, both written and oral (for the non-literate) informed consent was obtained from all participants in the study.
Subject Selection
Purposive random sampling of consenting 384 adult patients, aged 18 years+, receiving treatment at the selected health facilities during the study period of February to October 2023, and with visible clinical signs of onychomycosis, including discoloration, and with thickened nail textures was carried out.
Sample Collection
Nails of all participants were clinically examined. The fingernails and/or toenails were disinfected by applying 70% ethyl alcohol before sample collection to avoid contamination (Cheesbrough, 2000). Nail scrapings and trimmings were done with sterile surgical blades to scrape and trim small portions of the affected nail plate into a sterile polythene envelope, sealed and properly labelled. Samples were subsequently analyzed.
Preparation of Culture Media
Sabouraud Dextrose Agar (SDA) was prepared according to the manufacturer’s specifications. Five (5) mls of (what strength?) ciprofloxacin was added into (what quantity of medium?) the medium using a sterile syringe to inhibit the growth of bacteria and other contaminants.
METHODS
Each sample was divided into two parts: one for direct microscopy and the other for fungal culture. The nail samples were soaked in cryovate tubes containing 1ml each of potassium hydroxide (KOH) and agitated on a vortex mixer machine. A drop was then measured using a pipette onto a sterile slide. A drop of lactophenol cotton blue was added, covered with a clean slip and allowed to stand at room temperature until the material had cleared, and viewed using x40 and x10 magnification for the presence of filamentous septate hyphae, chlamydospores, yeast cells and macro or micro-conidia.
Scrapings from nail samples were also cultured using the pour plate method on SDA in sterile Petri dishes after inserting the nail. The plates were kept at room temperature (about 30oC) for 5 to 21 days, and observed periodically for growth, colony, colour, form, relief, consistency and morphology for identification.
Morphological Characterization
Isolated fungi were identified by comparing photographic Atlas and based on physiological and morphological properties (De Hoog, 2020). Colonies on SDA were classified according to shape, colour, size and margin characteristics. Plates with no visible growth after 21 days were considered negative. Visible colonies were stored in tween 80 broth and afterwards on SDA broth for molecular identification.
DNA Extraction
Eppendorf tubes containing frozen broth of fungal specimen from the refrigerator, allowed to thaw, then was centrifuged at 1400 rpm for 5 minutes to separate the broth from the pellet containing the fungal cells. About 500µl of previously prepared buffer was dispensed in each Eppendorf tube, and 200µl of the fungi broth was added to each. It was vortexed vigorously to separate the protein pellet from the DNA (supernatant) after which the DNA was harvested using sterile micro-pipettes into new well-labeled sterile Eppendorf tubes. The precipitate was transferred into a tube, washed with cold 70% ethanol and air-dried. Finally, the dried precipitate, containing genomic DNA, was mixed with 50 μl of Phosphate-buffered saline and stored at -20°C until use.
DNA Quantification and Gel Electrophoresis
Agarose Gel was prepared as recommended by manufacturer, cooled to 60oC, and 5µl of ethidium bromide added and mixed thoroughly. Combs were inserted into the gel tray and the agarose poured into the tank to and allowed to solidify. The edges of the tray were removed and the agarose in the tray was placed in the electrophoresis tank, more quantity of the TBE solution was poured into the tank to completely immerse the gel. Thereafter the well-forming combs were removed. Approximately 15µl each of the extracted DNA samples was loaded into the wells using a sterile pipette. 10µl of 100bp DNA ladder was loaded in one of the wells, connected to an electric source, the voltage set at 100V for 30 minutes and run. The resulting gels were viewed under a UV transilluminator.
PCR products were analyzed by running them on fresh agarose gel following the gel electrophoresis method above. Table 1 summarizes the primer sequences used for 25s and 18s rDNA amplification.
PCR typing were run as size markers to assist in the scoring of simple PCR types 1 through 3. The designation of complex PCR types was made by visual comparison of the separate pattern types run contemporaneously on agarose gels. Multiple comparisons were made to differentiate similar patterns with minor band variations. Samples were amplified de novo for each comparative gel, and all bands, including minor ones, were reproducible.
The data collected were analyzed using descriptive statistics to determine the prevalence of onychomycosis.
Table 1: Primers Used for PCR Amplification of 25S and 18S rDNA
| Primer | Primer Sequence | Amplicon Size (bp) |
| PRIMERSET | ||
| SQLE Primer for Mutation | F: 5’-GGGGCCTGGAGCTTATAGAT
R: 3’-GATCGAATAATGCCGGACAG |
702 |
| Single Nucleotide polymorphism (SNPS) 1R | F: 5’-CCACCAGCGGCTAATATAGAGTAA-3’(TTA>TTT: Leu393>Phe393) | |
| Single Nucleotide polymorphism (SNPS) 2R | F: 5’-CACCAGCGGCTAATATAGAGTATG-3/(TTA>TCA: Leu393>Ser393) | 702 |
| Single Nucleotide polymorphism (SNPS) 3R | F: 5’-TACCACCAGCGGCGAG-3’
/(TTC>CTC: Phe397>Leu397) |
702 |
| Single Nucleotide polymorphism (SNPS) 5R | F: 5/-TACCACCAGCGGCGAC-3’
|
702 |
| Strain Typing (TRNTSF2) | F: 5’-ACCGTATTAAGCTAGCGCTG’
(TTC>ATC: Phe397>Ile397) |
834 |
| Strain Typing (TRNTSR4) | R: 3’-TGCCACTTCGATTAGGAGGC’ | 834 |
| Strain Typing (TRNTSC1) | F: 5’-CGAGACCACGTGATACATGCG’ | 502 |
| Strain Typing (TRNTSR1) | R: 3’-CTCAGTCGAACCGTGAGGC’ | 502 |
RESULTS
Table1 shows that among the 384 samples collected, 210 were positive by direct microscopy technique, while 195 (92.86%) were culture positive. The false negative by direct microscopy were 15 (7.14%).
Out of 226 fungal species isolated through culture, 159 (70.36%) were T. rubrum, followed by Aspergillus spp. 30 (13.27%), Candida (Yeast) 19 (8.41%), T. interdigitale 11 (4.87%) and E. floccosum 8 (3.54%). Table 2. The table further shows that on ycomycosis infection was very high in toenails; 210(68.48%), followed by infection on both toes and fingernails; 8 (24.24). Infection on fingernails was 8 (18.18%).
Table 2: Results of Direct Microscopy and Culture
| Direct Microscopy | Culture Positive (%) | Culture Negative (%) | Total (%) |
| Positive | 195 (92.86%) | 15 (7.14%) | 210 (100%) |
| Negative | 31 (17.82%) | 143 (82.18%) | 174 (100%) |
| Total | 226 (58.85%) | 158 (41.15%) | 384 (100%) |
Table 3: Occurrence of Fungi Species Isolated from the Nails on Cultural Plates
| Species | Site of Infection | Frequency | Percentages | ||
| Toenails | Fingernails | Toes/Fingernails | |||
| T. rubrum | 154 | 2 | 3 | 159 | 70.36 |
| Aspergillus spp. | 23 | 2 | 4 | 30 | 13.27 |
| Candida spp. | 15 | 3 | 1 | 19 | 8.41 |
| T. interdigitale | 10 | 1 | 0 | 11 | 4.87 |
| E. floccosum | 8 | 0 | 0 | 8 | 3.54 |
| Total | 210 | 8 | 8 | 226 | 100 |
Plate 1 shows that the Squalene Epoxidase (SQLE) gene was successfully amplified in all samples, regardless of the presence of mutations. Mutations in the SQLE gene did not affect the amplification process which appeared consistent after analysis using 1.5% agarose gel electrophoresis. SQLE primer was used for the identification of Trichophyton rubrum isolates. Lane (L) is the 100bp DNA ladder, and Lanes 1-12 are the Trichophyton rubrum amplicons which showed intense bands with varying molecular weights. The amplicons for the positive samples were all visible at the 720bp DNA markers.

Plate 1: Amplification of SQLE gene common to all samples regardless of mutations on Agarose gel electrophoresis
Strain Typing of T. Rubrum by Specific Amplification of the TRS-1 and TRS-2 Repeat Regions
Primers TrNTSF-2 and TrNTSR-4 were used to amplify the TRS-1 repeat regions from seven T. rubrum strains of rDNA types 1 through 7. The major fragment amplified from each rDNA type showed stable length variation equivalent to one TRS-1 repeat unit, except the variation in length between types 2 and 7. A ladder of minor bands ranging in size from 500 to 550 bp and stepping up in increments of 600 bp was also present. These simple patterns were designated PCR types 1 through 7 (Plate 2a; lanes 1 through 7) and intense bands were visible at 500bp to 600bp (Plate 2b; lanes 8 through 12).
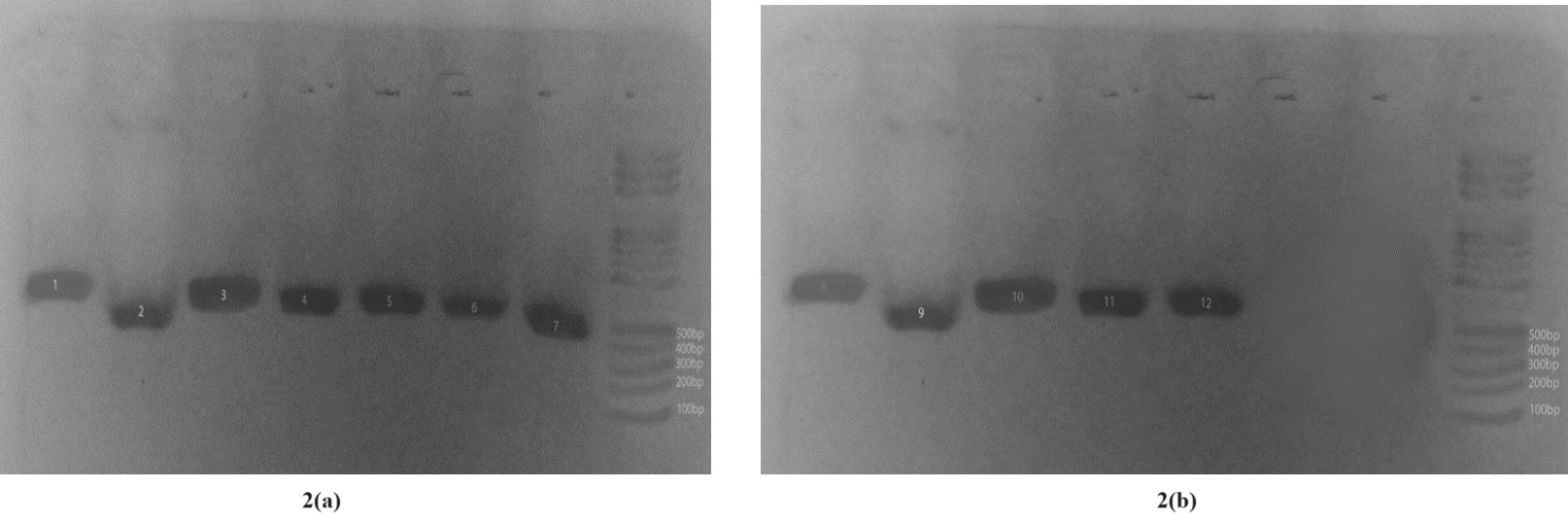
Plate 2a: TRS-1 Amplification; samples 1-7. Plate 2b: TRS-1 Amplification; samples 8-12
The non-transcribed Spacer (NTS) region containing TRS-2 was amplified using primers TrNTSR-1 and TrNTSC-1. A single 502-bp product (lane 12), representing two complete copies of TRS-2, was amplified. Variation in the TRS-2 fragment occurred mostly in strains with a single copy of TRS-1. Lanes 6-12 were the only positive isolates (Plate 3).

Plate 3: TRS-1 Amplification; positive samples are 6 to 12
DISCUSSION
Results showed 210 positive samples by direct microscopy technique, while 195 (92.86%) were culture positive. The false negatives by direct microscopy were 15 (7.14%). These findings are as reported by Wijesuriya et al. (2014) that in patients who were clinically suspected of having superficial fungal foot infections, 14% showed negative culture results. The differences in the direct microscopy and culture results may be due to the use of antifungal ointments by some individuals from who samples were collected. This might have inhibited the growth of some fungal species.
In this study, the finding of high isolation rates of T. rubrum contradicts results of Chukwuka et al. (2013) that Cladosporium spp. had the highest occurrence (64.0%). Similarly, findings of Baki et al. (2018) also varied with present findings. The predominant (37.43%) pathogen in their study were yeast species. Yeasts were closely followed by T. rubrum (33.30%), while moulds were 29.12% of cases. These variations may be due to study location and sites of sample collection. Trichophyton spp. are known to be more prevalent in toenails.
Attempts to identify strains in the dermatophyte fungus Trichophyton rubrum have been made using various molecular typing techniques. Previous investigations primarily utilized random amplified polymorphic-DNA methods, which typically resulted in the identification of a single strain or a limited number of strain lineages. (Gra¨ser et al.,1999). Random amplification of polymorphic DNA is useful for the differentiation of several anthropophilic dermatophytes. Mycoses 40:405–409) A prior method, involving the analysis of restriction fragment length polymorphisms in the non-transcribed region of the rDNA spacer, was introduced for strain differentiation in this species (Jackson et al., 1999). In the current study, these polymorphisms were not only confirmed, but the existence of two new sub-repeat elements, TRS-1 and TRS-2, in the NTS region were also revealed. Additionally, the study demonstrated that variations in the copy number of these elements contribute to the previously observed NTS length polymorphisms. The specific amplification of NTS regions containing these elements emerge as a straightforward, swift, and discriminative method for strain typing of T. rubrum.
Varieties of these common PCR types may exhibit specific features related to their ability to cause onychomycosis disease or their capacity to infect, which have contributed to their widespread dissemination (Gra¨ser et al.,1999). The considerable diversity found in the rDNA NTS stands in contrast to a recent description of T. rubrum being strictly clonal. The fluctuations in the NTS sub-repeat copy numbers likely stem from the unequal exchange of sister chromatids during mitosis (Szostak and Wu, 1980). Polymorphisms in repetitive DNA segments resulting from uneven crossing-over are confined to specific locations and were associated with variations due to recombination and mutation elsewhere in the genome.
Findings of this study have motivated exploration of alternative cryptic, polymorphic repetitive sequences to distinguish between strains of T. rubrum. Identifying additional unlinked variable markers would enable allelic analysis using existing population genetic techniques (Taylor et al., 1999). The development of Allele-specific PCR method facilitates allele analysis of clinically significant mutations, ensuring reliable discrimination between two highly distinct alleles.
CONCLUSION
The study presented findings on onychomycosis, focusing on molecular typing techniques for Trichophyton rubrum, and revealed the presence of new sub-repeat elements in the non-transcribed region of the rDNA spacer. This information contributes to the understanding of genetic diversity and strain differentiation in T. rubrum.
The study also provides valuable insights into the prevalence and associated factors of onychomycosis, emphasizing the need for a holistic understanding that encompasses a molecular perspective.
RECOMMENDATIONS
The variation in predominant fungal species isolated from toenail and fingernail samples highlight the importance of considering geographical and environmental factors. It is recommended to conduct further studies to understand regional variations in fungal species prevalence.
Furthermore, the study on the strain typing of Trichophyton rubrum using molecular techniques provides valuable insights. Further research and adoption of advanced molecular typing methods can enhance our understanding of infecting onychomycosis fungal strains.
REFERENCES
- Adejuwon, J. O. (2012). Rainfall seasonality in the Niger Delta belt, Nigeria. Journal of Geography and Regional Planning, 5(2), 51.
- Agada, S., and Nirupama, N. (2015). A serious flooding event in Nigeria in 2012 with specific focus on Benue State: a brief review. Natural Hazards, 77; 1405-1414.
- Baki, A.S., Bello, A., Mohammad, U.K., Farouq, A.A., Usman, A.A., et al. (2018) A Survey for Onychomycosis among Patients Attending General Hospital Ammannawa, Sokoto State Nigeria. Journal Microbiology Pathology 2: 108.
- Cheesbrough, M. (2000). District laboratory practice in tropical countries, Part 2. Cambridge University Press.
- Chukwuka, K. S., Iwuagwu, M. I. and Uka, U.N. (2013). Evaluation of nutritional components of Caricapapaya At different stages of ripening. IOSR Journal of Pharmacy and Biological Sciences 6(4): 13-16
- de Berker, D. A. R., André, J. and Baran, R. (2007). Nail biology and nail science. International Journal of Cosmetic Science, 29(4); 241–275.
- Gra¨ser, Y., Ku¨hnisch, J. and Presber, W., (1999). Molecular markers reveal exclusively clonal reproduction in Trichophyton rubrum. Journal of Clinical Microbiology. 37:37 13–3717.
- Jackson, C. J., Barton, R. C. and Evans, E. G. V. (1999). Species identification and strain differentiation of dermatophyte fungi by analysis of ribosomal- DNA spacer regions. Journal of Clinical Microbiology. 37:931–936.
- Jazdarehee, A., Malekafzali, L., Lee, J., Lewis, R. and Mukovozov, I. (2022). Transmission of Onychomycosis and Dermatophytosis between Household Members: A Scoping Review. Journal of Fungi, 8, 60.
- Szostak, J., and Wu, R. (1980). Unequal crossing over in the ribosomal DNA of Saccharomyces cerevisiae. Nature 284:426–430.
- Taylor, J. W., Geiser, D. M., Burt, A. and Koufopanou, V. (1999). The evolutionary biology and population genetics underlying fungal strain typing. Microbiology Review. 12:126–146.
- Westerberg, D. P. and Voyack, M. J. (2013). Onychomycosis: Current trends in diagnosis and treatment. American Family Physician, 88(11); 762–770.
- Yenişehirli, G., Karat, E., Bulut, Y. and Savcı, U. (2012) Dermatophytes Isolated from the Mosques in Tokat, Turkey. Mycopathologia, 174, 327–330.