
Density and Phenotypic Diversity of Arbuscular Mycorrhizal Fungi in a Revegetated Tailing Storage Facility in Ghana
- Ernest Kofi Amankwa Afrifa
- Abdul-Rahaman Afitiri
- Jennifer Badu
- 728-743
- Jan 17, 2024
- Education
Density and Phenotypic Diversity of Arbuscular Mycorrhizal Fungi in a Revegetated Tailing Storage Facility in Ghana
Ernest Kofi Amankwa Afrifa*1, Abdul-Rahaman Afitiri2, Jennifer Badu2
Department of Environmental Science, School of Biological Sciences, University of Cape Coast, Cape Coast, Ghana.1
Institute of Environmental Technology and Process Engineering, Chair of Biotechnology of Water Treatment, Brandenburg University of Technology Cottbus-Senftenberg, 03046 Cottbus, Germany2
*Corresponding Author
DOI: https://doi.org/10.51244/IJRSI.2023.1012056
Received: 27 November 2023; Revised: 15 December 2023; Accepted: 20 December 2023; Published: 16 January 2024
ABSTRACT
Arbuscular Mycorrhizal fungi (AMF) aid the establishment of vegetation on stressed soils as they contribute to soil ecological functions as well as improvement in plant growth. Understanding the mycorrhizal diversity and density within an area is an important step in harnessing the immense benefits of mycorrhiza. This study assessed the diversity and density of AMF in a revegetated tailing storage facility in Ghana. A balanced hierarchical sampling scheme with 4 stages and 72 sampling points was used over four sampling periods. Seventeen AMF spore types from nine genera (Acaulospora, Claroideoglomus, Diversispora, Gigaspora, Glomus, Racocetra, Rhizophagus, Septoglomus and Scutellospora) and five families (Acaulosporaceae, Claroideoglomeraceae, Diversisporaceae, Gigasporaceae and Glomeraceae) were isolated and identified from the samples. The density of spores varied between 24 and 817; 21 and 303; 21 and 321; and 24 and 875 spores 100 g-1 dry tailing solids respectively for the four sampled periods. The spore densities for the control samples also varied between 85 and 307; 65 and 248; 90 and 321 and 70 and 287 spores 100 g-1 dry soil for the respective sampling periods. Recorded spore densities varied significantly within sampling locations (p = 0.000) and between sampling periods (p = 0.000). The density and diversity of the AMF found in this study were relatively high and may have contributed significantly to the revegetation of the storage facility. These findings are important pointers for the adoption of mycorrhization in vegetation establishment and ultimately rehabilitation of decommissioned tailings storage facilities in Ghana. Further studies should however be done to determine how the various vegetation types found on the TSF affect AMF diversity and density.
Key words: Arbuscular mycorrhizal fungi, tailing storage facility, mining, vegetation, Ghana
INTRODUCTION
Gold mining in Ghana continues to grow in size and significance (McQuilken & Hilson, 2016) as the sector continues to play a significant role in the socioeconomic development and livelihoods of over 15 million people in the country (Akabzaa, 2009); and together with other minerals extracted in the country contributes significantly to national development (Ros–Tonen et al., 2021). Ghana’s mineral potential and the country’s contribution to global minerals output, especially gold is well acknowledged (Ofosu et al., 2020; Essah, 2021; Yiridomoh, 2021). On the average, mining accounts for about a third of gross foreign exchange and about 7% of gross domestic product. Its contribution to government tax revenue is around 4%, employing about 0.7% of the working age population (Baah, 2005; ICMM, 2015). Several studies however, have reported the negative impacts of mining activities on the environment, and the microbial diversity and activity in soils (Aghetara, 2012; Yankson & Gough, 2019). High-volume toxic waste material (tailing) is generated from mining projects and their management and disposal informs if a mining project is environmentally friendly or not (Kemp et al., 2021).
Mining, mineral processing, and metallurgical extraction result in the production of a high volume of unwanted material in the form of overburden, rock waste, tailings, spoils, sludge and process water. The vast majority of the mined material ends up as tailings which, represent the most voluminous waste at metal mine sites especially in instances where low-grade ores are utilized (Lottermoser, 2007). Tailings are the materials remaining after extraction and beneficiation of ores and are characterized by elevated concentrations (1–50 g kg-1) of metals such as arsenic, cadmium, copper, manganese, lead, and zinc (Boulet & Larocque, 1998). With no organic matter or macronutrients, and usually exhibiting acidic pH, although some tailings may be alkaline (Krzaklewski & Pietrzykowski, 2002; Titshall et al. 2013), tailings remain without normal soil structure and generally support a severely stressed heterotrophic microbial community (Mendez et al., 2007). Mine tailings are usually stored in surface containments within embankments, usually referred to as tailings storage facilities (TSFs) or tailings dams. These storage facilities or dams may range from a few hectares to thousands of hectares in size and because of this, they leave the largest “footprint” of any mining activity on the landscape (Lottermoser, 2007). The environmental impact of mine tailing disposal sites is enormous, as unreclaimed mining sites generally remain unvegetated for years, and exposed tailings can spread over tens of hectares via eolian dispersion and water erosion (González & González-Chávez, 2006).
The key long-term goal of tailing disposal and management is to prevent the mobilization and release into the environment of toxic constituents of the tailing. In most instances, these tailing facilities are revegetated and put under agricultural usage (Prayudyaningsih et al., 2019). The practice of appropriate revegetation techniques in mine tailing is necessary to reduce erosion, protect soil against degradation, and limit the spread of metal contamination (Mendez & Maier, 2008; Carrasco et al., 2010). Soil microorganisms are known to play a key role in the mobilization and immobilization of metals thereby changing their availability to plants (Wang et al., 2022).
Root-colonizing symbiotic microorganisms such as arbuscular mycorrhizal fungi (AMF) are mainly involved in phytoremediation (Cicatelli et al., 2014) and can be used as a complement to immobilization strategies (Gaur & Adholeya, 2004) especially for heavy metals. AMF are soil microorganisms that establish mutual symbioses with the majority of higher plants, providing a direct physical link between soil and plant roots (Kumar & Singh, 2019). They belong to the phylum Glomeromycota and are ubiquitous components of the soil microbiome that form nutritional associations with a great majority of plants in various ecosystems (Smith & Read, 2010). AMF occur in almost all habitats and climates (Davison et al., 2011) including disturbed soils such as those derived from mining activities (Johnson, 2010; Prayudyaningsih et al., 2019). However, soil degradation brings about changes in the diversity and abundance of AMF populations (Lekberg & Koide, 2005; An et al., 2008; Silva et al., 2022). An important function of AMF is the provision of mineral nutrients to plants in exchange for photosynthates and aiding in regulation and maintenance of plant productivity and diversity (Johnson, 2010). Changes in AMF communities can strongly affect plant communities in either way (Klironomos et al., 2011).
AMF have been shown to aid the establishment of vegetation on stressed soils (Ardestani et al., 2019). The extent to which vegetation is established successfully on a nutrient-poor tailing storage facility will depend on the consortia of mycorrhiza present in a given area. Many researchers have reported the occurrence of various mycorrhiza in plant roots growing in heavy metal contaminated soils (Sharma & Sharma, 2013; Yang et al., 2017) whilst others have highlighted their importance (Gaur & Adholeya, 2004), however, very little work (Asmah, 1996) has been done on mycorrhiza in general in Ghana and none on abandoned tailing storage facilities in any of the mining companies in Ghana. The diversity of mycorrhizal species in Ghanaian soils and revegetated tailing storage facilities is not well known. Though a lot of decommissioned tailing storage facilities can be found in the mining areas of Ghana, expensive conventional engineering options are usually used in their rehabilitation, leaving out the cheaper option of the application of mycorrhiza. Thus, the wealth of benefits that can be derived from these organisms remain untapped due to the limited knowledge on them. Understanding the mycorrhizal diversity and density within an area is a first step in harnessing the immense benefits of mycorrhiza. It is on this basis that this study was carried out to assess the diversity and density of AMF in a revegetated tailing storage facility in Ghana.
MATERIALS AND METHODS
Study Area
The study was carried out within the concession of Abosso Goldfields Limited (AGL), a Ghanaian registered company, which operates the Damang Gold Mine. With a total concession area of the 52.39km2 (8111 ha), the Damang Mine is located about 1.5km North-east of the Damang village in the Prestea Huni-Valley Municipality of the Western Region of Ghana (Figure 1). It is also 45km North-East of Tarkwa, the capital of the Tarkwa-Nsuaem Municipality. It lies on latitude 5°’15’05″N and longitude 1°59’17″W.
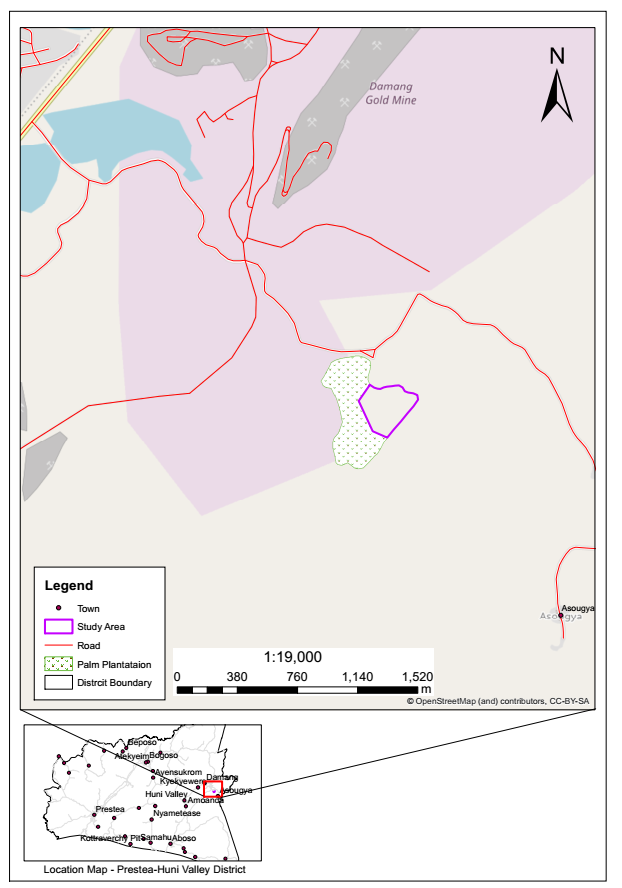
Figure 1: Map of study area
Sampling Design
A modified balanced hierarchical sampling scheme with four stages was adopted in this study following the works of Youden and Mehlich (1937). An area of approximately 150 m x 150 m was mapped out within the TSF as the experimental site. Nine (9) primary locations, designated as sampling stations, which were 75 m apart, were selected. At each station, two (2) substations were selected 29 m apart. Two (2) sampling areas 17 m apart were located at each substation. Finally, in each sampling area, two (2) sampling points, 5 m apart, were located. These distances were not rigidly adhered to and this, combined with the complete freedom to direction, permitted the exercise of the utmost discrimination in securing the samples under comparable conditions. In all, 72 sampling points were used in this study as indicated in Table 1 with the schematic presentation of the sampling design shown in Figure 2. The study was conducted over four sampling periods (September – November 2020; December 2020 – February 2021; March – May 2021 and July – August 2021).
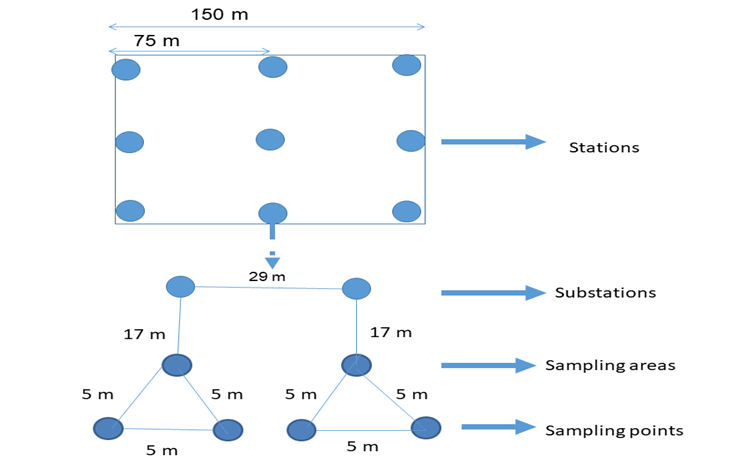
Figure 2: Schematic representation of sampling design [Adopted from Youden and Mehlich (1937)].
Sample Collection
Approximately 2 kg of tailing were collected up to a depth of 20cm using a plastic trowel from 8 sampling points per sampling station. The samples were transported to the laboratory in labeled plastic sample bags. The labeling and identification of the samples were facilitated by the adoption of a numbering system where each sample was codified in the x-y plane. The nine stations were given the numbers 0 to 8 and appear in the x- position of the code. The eight sample points at each station were numbered 1 to 8 and occupied the y-position in the number. In all, 288 tailing samples were collected over the sampling period. Samples were also collected from an unmined location to serve as a control.
Preliminary Sample Preparation
The tailing samples were air-dried and sieved with a 2mm sieve to remove coarse materials such as stones and other debris. Tailing samples were stored at room temperature for further laboratory analysis.
Spore Isolation and Identification
All tailing samples were analysed for their AM fungi spore content by modification of methods for wet sieving according to Gerdemann and Nicolson (1963) followed by sucrose centrifugation (Oehl et al. 2003). 100g each of the previously sieved tailing samples were suspended in 400 ml of tap water and vigorously washed for 10 minutes to free the spores from aggregates of soil or organic materials. Calgon was not added as the samples were not clayey. The tailing suspension was then wet-sieved through a series of sieves of three different pore sizes, 750µm, 350µm and 45µm. The wet sieving was done by washing the suspension in lukewarm running tap water. Roots and other coarse debris collected on the coarse (750-µm) sieve, while spores were captured on the finer sieves. The contents from the two sieves with the smallest pore sizes were pooled and centrifuged at 1750 rpm for 5 minutes in a horizontal rotor. The supernatant liquid was carefully decanted, the pellet resuspended in 60% sucrose solution and further centrifuged for 5 minutes. The sucrose fraction containing AM fungal spores was then sieved through a 30µm sieve and rinsed thoroughly with running water to remove the sugar. The sieving was transferred onto petri dishes, counted under a compound microscope and the number of spores in the dish expressed as spores/g of the tailing sample. The sievings that were not immediately observed were then preserved in Ringer’s solution (0.6g NaCl, 0.01g CaCl2, 0.01g KCl and 0.01 MgCl2 in 100ml distilled water) for later analysis.
For AM spore identification, the spores were mounted on microscope slides as described by Schenck & Perez (1990) and identified to the level of genus or species. Identifications were based on current species descriptions and identification manuals (Schenck & Perez, 1990; INVAM, 2014).
The morphological variables used for spore identification, as prescribed by de la Cruz (1991), were: occurrence of sporocarp, its shape, colour and size; occurrence of peridium and its characteristics; colour of spores, their size and shape; number of spore walls, their colour, thickness and ornamentation; and attachment of spores, shape and type of occlusions.
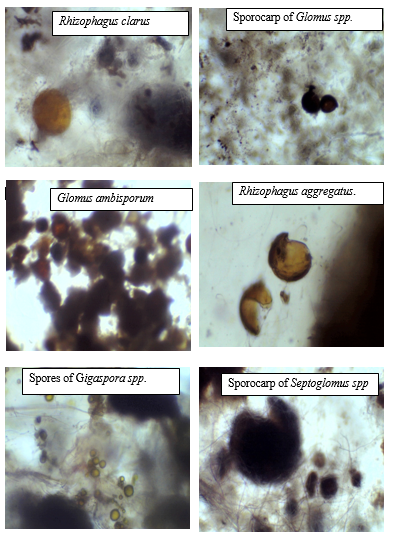
Figure 3: Samples of isolated AMF spores
Data analyses
Diversity and Dispersion
Diversity and dispersion of the AMF were evaluated using both the Simpson’s diversity index (D) and Shannon-Wiener index (H’).
Simpson’s Diversity index (D)
The Simpson’s diversity index was computed through the equation

Where n is the number of each AMF species and N the total number of individuals of all species.
Shannon-Wiener Index (H’)
The index was calculated using the equation
![]()
Where s is the total number of species and p is the relative abundance of the ith species.
Statistical analysis
Conventional statistical analysis was performed using the IBM SPSS 24.0 software (SPSS Inc., Chicago, IL, USA). Multiple comparisons and analyses of variance (ANOVA) were used to determine the differences among and within spore densities and sampling periods for both tailing and control samples. Differences at p< 0.05 were considered significant.
RESULTS
Arbuscular Mycorrhizae Fungal Spore Density in the Soil
The AMF spore density (spores 100 g-1 dry tailing solids) at the various sampled locations are shown in Table 1 below.
Table 1: Mean AMF spore density (100 g-1 dry soil)
| SP1 | SP2 | SP3 | SP4 | P-value | ||
| Sampling periods | Locations | |||||
| Tailings | 0.000 | 0.000 | ||||
| Low | 21 | 21 | 21 | 24 | ||
| High | 817 | 303 | 321 | 875 | ||
| Mean | 253±201a | 115 ±32b | 119 ±52b | 262±204a | ||
| Control | 0.053 | 0.000 | ||||
| Low | 85 | 65 | 90 | 70 | ||
| High | 307 | 248 | 321 | 287 | ||
| Mean | 165±76.45a | 152±64a | 167±82a | 151±74a | ||
Means followed by similar letters are not statistically significant. SP denotes Sampling period
AMF spores were present in all tailing samples for all sampled periods. As shown in Table 1 above, the density of spores varied between 24 and 817; 21 and 303; 21 and 321; and 24 and 875 spores 100 g-1 dry tailing solids respectively for the four sampled periods. The spore densities for the control also varied between 85 and 307; 65 and 248; 90 and 321 and 70 and 287 spores 100 g-1 dry soil for the respective sampling periods. The highest spore density of 875 spores 100 g-1 dry tailing was obtained during the fourth sampling period with the lowest of 21 spores 100 g-1 dry tailing being recorded during the first and third sampling periods. The fourth sampling period returned the highest densities, with a mean of 262.17±204.4 spores 100 g-1 dry tailing whilst the second period returned the least densities with an average of 114.72±32 spores 100 g-1 dry tailing. Statistically significant differences existed between the sampled tailing locations (p = 0.000) and across the sampled periods (p = 0.000). Statistically significant differences were also observed between the densities at the various control points (p = 0.000); however, the differences across the sampled periods were not statistically significant (p = 0.053). Between the tailings and control locations, no significant differences existed between the various sampled points (p = 0.536) and across the sampled periods (p = 0.510).
Diversity of Arbuscular Mycorrhizae Fungi
Diversity of AMF in analysed tailing and control (soil) samples are respectively shown in Tables 2 and 3. Seventeen AMF spore types from nine genera (Acaulospora, Claroideoglomus, Diversispora, Gigaspora, Glomus, Racocetra, Rhizophagus, Septoglomus and Scutellospora) and five families (Acaulosporaceae, Claroideoglomeraceae, Diversisporaceae, Gigasporaceae and Glomeraceae) were isolated and identified from the tailing samples (Table 2). Out of the seventeen spore types, four belonged to the genus Glomus, three to the genus Acaulospora, and two each to Gigaspora, Rhizophagus and Septoglomus whilst Claroideoglomus, Diversispora, Racocetra and Scutellospora had one each. Within the genus Glomus, one spore type could not be identified to the species level. All tailing samples contained at least 2 spore types. The dominant spore type within the tailing samples was Glomus ambisporum, which accounted for 23.5% of the total spores isolated. The least was the unidentified Glomus sps that accounted for only 0.063% of the isolated spores. Four species, Diversispora epigea, Gigaspora margarita, Glomus cerebriformes and Scutellospora calospora were not isolated from any of the locations during the second and third sampling periods. The mean numbers of the isolated spores are shown in Figure 4.
Table 2: Diversity of AMF spores in tailing samples
| Spore type | SP1 | SP2 | SP3 | SP4 | Total | Rank |
| Acaulospora denticulate | 1913 | 117 | 154 | 2062 | 4246 | 7 |
| Acaulospora foveata | 673 | 814 | 874 | 716 | 3077 | 9 |
| Acaulospora tuberculata | 1285 | 311 | 342 | 1280 | 3218 | 8 |
| Claroideoglomus luteum | 1256 | 68 | 129 | 1329 | 2782 | 10 |
| Diversispora epigea | 370 | 0 | 0 | 387 | 757 | 13 |
| Gigaspora decipiens | 2071 | 94 | 157 | 2155 | 4477 | 6 |
| Gigaspora margarita | 18 | 0 | 0 | 28 | 46 | 16 |
| Glomus ambisporum | 4456 | 1963 | 1959 | 4284 | 12662 | 1 |
| Glomus cerebriformes | 18 | 0 | 0 | 143 | 161 | 14 |
| Glomus marcocarpum | 1903 | 611 | 624 | 1933 | 5071 | 4 |
| Glomus sps(Sporocarpia) | 16 | 1 | 1 | 16 | 34 | 17 |
| Racocetra gregaria | 635 | 46 | 46 | 685 | 1412 | 11 |
| Rhizophagus aggregatus | 1188 | 1395 | 1417 | 1226 | 5226 | 2 |
| Rhizophagus clarus | 1326 | 1229 | 1242 | 1424 | 5221 | 3 |
| Septoglomus constrictum | 188 | 148 | 153 | 277 | 766 | 12 |
| Septoglomus deserticola | 849 | 1463 | 1472 | 886 | 4670 | 5 |
| Scutellospora calospora | 45 | 0 | 0 | 45 | 90 | 15 |
Figure 4: Mean number (to the nearest whole) of spores for tailing samples
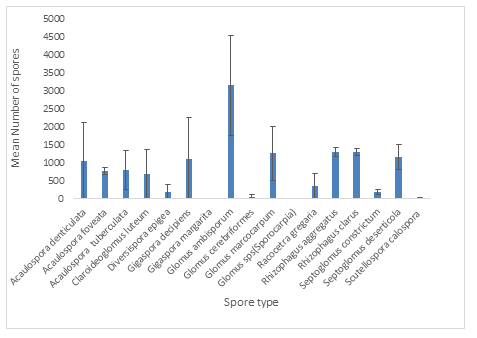
Table 3: Diversity of AMF spores in control samples
| Spore type | SP1 | SP2 | SP3 | SP4 | Total | Rank |
| Acaulospora tuberculata | 140 | 167 | 128 | 131 | 566 | 3 |
| Claroideoglomus luteum | 64 | 107 | 74 | 86 | 331 | 6 |
| Dentiscutata nigra | 480 | 473 | 476 | 495 | 1924 | 1 |
| Gigaspora decipiens | 50 | 66 | 74 | 81 | 271 | 7 |
| Glomus ambisporum | 370 | 378 | 378 | 334 | 1460 | 2 |
| Glomus marcocarpum | 100 | 116 | 127 | 119 | 462 | 5 |
| Rhizophagus aggregatus | 97 | 141 | 108 | 119 | 465 | 4 |
| Septoglomus constrictum | 17 | 30 | 42 | 27 | 116 | 8 |
Eight AMF spore types from seven genera (Acaulospora, Claroideoglomus, Dentiscutata, Gigaspora, Glomus, Rhizophagus and Septoglomus) and four families (Acaulosporaceae, Claroideoglomeraceae, Gigasporaceae and Glomeraceae) were isolated and identified from the tailing samples (Table 3). Of the eight spore types identified, two belonged to the genus Glomus whilst Acaulospora, Claroideoglomus, Dentiscutata, Gigaspora, Rhizophagus and Septoglomus had one each. All control samples contained at least two spore types. The dominant spore type within the tailing samples was Dentiscutata nigra, which accounted for 34% of the total spores isolated. The least was Septoglomus constrictum which accounted for only 2% of the isolated spores.
Quantitative Diversity Indices of AMF Spore Species in the Tailing Samples
Species Diversity and Dispersion
The diversity and dispersion of the AMF within the TSF are represented by the indices in Tables 4 and 5.
Table 4: Mean Shannon-Wiener Diversity Indices (H’) for the sampling points
| Sampling Stations | Sampling points | |||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | |
| 0 | 1.54±0.29 | 1.94±0.24 | 1.43±0.43 | 1.39±0.31 | 1.50±0.29 | 1.53±0.13 | 1.64±0.24 | 1.59 ±0.21 |
| 1 | 1.38±0.09 | 1.44±0.2 | 1.63±0.05 | 1.48±0.00 | 1.28±0.14 | 1.32±0.11 | 1.28±0.04 | 1.55±0.07 |
| 2 | 1.26±0.05 | 1.55±0.06 | 1.38±0.07 | 1.62±0.02 | 1.58±0.03 | 1.30±0.08 | 1.64±0.07 | 1.55±0.09 |
| 3 | 1.55±0.19 | 1.57±0.13 | 1.52±0.16 | 1.35±0.05 | 1.27±0.15 | 1.56±0.15 | 1.54±0.37 | 1.54±0.24 |
| 4 | 1.68±0.42 | 1.61±0.2 | 1.19±0.04 | 1.58±0.10 | 1.84±0.05 | 1.79±0.15 | 1.66±0.08 | 1.62±0.05 |
| 5 | 1.30±0.24 | 1.16±0.19 | 1.56±0.19 | 1.14±0.14 | 1.31±0.12 | 1.47±0.07 | 1.67±0.02 | 1.69±0.03 |
| 6 | 1.89±0.00 | 1.66±0.01 | 1.44±0.19 | 1.70±0.28 | 1.76±0.40 | 1.65±0.44 | 1.28±0.36 | 1.70±0.19 |
| 7 | 1.50±0.01 | 1.33±0.17 | 1.55±0.11 | 1.21±0.19 | 1.16±0.05 | 1.40±0.16 | 1.42±0.05 | 1.56±0.03 |
| 8 | 1.66±0.14 | 1.65±0.05 | 1.22±0.05 | 1.56±0.02 | 1.64±0.09 | 1.25±0.29 | 1.33±0.13 | 0.99±0.3 |
| C | 1.61±0.17 | 1.59±0.07 | 1.77±0.05 | 1.58±0.02 | 1.70±0.19 | 1.45±0.35 | 1.26±0.01 | 1.16±0.01 |
*Rows = Sampling stations; Columns = sampling points; C = control
Table 4: Mean Simpson’s Index of Diversity (D’) for the sampling points
| Sampling
Stations |
Sampling points | |||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | |
| 0 | 0.69±0.08 | 0.86±0.05 | 0.66±0.14 | 0.67±0.12 | 0.75±0.13 | 0.77±0.04 | 0.79±0.06 | 0.76±0.06 |
| 1 | 0.76±0.03 | 0.75±0.05 | 0.81±0.02 | 0.76±0.04 | 0.78±0.01 | 0.73±0.03 | 0.73±0.08 | 0.82±0.03 |
| 2 | 0.72±0.04 | 0.83±0.01 | 0.78±0.07 | 0.79±0.05 | 0.81±0.01 | 0.70±0.09 | 0.82±0.01 | 0.77±0.07 |
| 3 | 0.77±0.08 | 0.79±0.05 | 0.78±0.08 | 0.65±0.01 | 0.68±0.07 | 0.81±0.01 | 0.79±0.05 | 0.78±0.03 |
| 4 | 0.82±0.06 | 0.81±0.07 | 0.69±0.07 | 0.79±0.06 | 0.84±0.00 | 0.85±0.01 | 0.82±0.05 | 0.82±0.02 |
| 5 | 0.74±0.02 | 0.68±0.07 | 0.77±0.04 | 0.62±0.08 | 0.72±0.10 | 0.80±0.03 | 0.81±0.03 | 0.84±0.02 |
| 6 | 0.83±0.01 | 0.80±0.01 | 0.69±0.06 | 0.79±0.02 | 0.82±0.04 | 0.77±0.06 | 0.69±0.05 | 0.80±0.00 |
| 7 | 0.78±0.01 | 0.78±0.01 | 0.76±0.04 | 0.67±0.04 | 0.64±0.05 | 0.73±0.09 | 0.77±0.02 | 0.78±0.05 |
| 8 | 0.80±0.03 | 0.80±0.01 | 0.71±0.01 | 0.76±0.02 | 0.79±0.01 | 0.67±0.12 | 0.71±0.08 | 0.60±0.08 |
| C | 0.76±0.03 | 0.75±0.02 | 0.81±0.05 | 0.75±0.06 | 0.78±0.04 | 0.74±0.05 | 0.65±0.03 | 0.64±0.02 |
*Rows = Sampling stations; Columns = sampling points; C = control
The mean Shannon-Weiner indices (H’), as seen from Table 4, were low to moderate for both tailing and control samples. The lowest mean H’ index for the tailing samples of 0.99±0.3 was recorded at Location 88 whereas the highest index of 1.94 occurred at Location 02. For the control samples, Location C3 recorded the highest H’ index of 1.77±0.05 with the lowest of 1.16±0.01 occurring at C8. Statistically significant differences (p = 0.049) existed in the mean Shannon-Wiener diversity Indices (H’) of the tailing samples between the sampling stations. However, the differences between the sampling points were not statistically significant (p = 0.882).
The mean Simpson’s Indices of Diversity for the sampling points are as shown in Table 5. Location 02 recorded the highest Simpson’s Index of Diversity of 0.86±0.05 with the lowest of 0.62±0.08 occurring at Location 54. For the control samples, C8 recorded the highest Simpson’s index of 0.81±0.05 while the lowest of 0.64±0.02 was recorded at C8. Statistically significant differences were found to exist between the Simpson’s indices at the sampling stations (p = 0.01) whereas no statistically significant differences existed between the indices at the sampling points (p = 0.531).
DISCUSSION
The results of this study revealed the presence of AMF spores in all the samples analysed, with more than 70% of the samples containing 100 spores 100 g-1 dry soil or more.
The relatively high spore densities identified in this study supports other studies by Zhao et al. (2001) and Dalpe and Aiken (1998) who found AMF at densities of 55 to 1,908 spores 100 g-1 soil and 70 spores 100 g-1 of dry soil respectively. Spore densities recorded in this study also showed significant variations within sampling locations and between the sampling periods, supporting the assertion of Zhao et al. (2003) that large variations in AMF spore densities associated with plant species (even same species) at different sites exit. These are attributable to variations in factors such as microclimate (Silva-Flores et al., 2019), management practices, nutrient levels, plant composition and edaphic factors such as pH, heavy metal levels and moisture content (Bever et al., 2001; Johnson et al., 2013). The variations in the number of isolated spores at the various sampling locations are thus expected as the factors influencing spore numbers vary even at the micro-scale in space and in time (Davison et al., 2011). The TSF, and more especially the study site had been put under different usages; some sections have been put under plantation cropping, with the dominant stands being oil palm. A mixed stand of oil palm and several uncultivated grass species also occupy sections of the study area while others are under leguminous trees and Vetiver grass cultivation; and thus, possesses different histories of management practices. The control area is also a forest stand composed of dense woody plants with minimal undergrowth. As the different plant stands will require different cultural management practices, localized spore densities will differ as it has been established that cultural practices affect the amounts of spores in the soil. Again, one of the means of spore transport is through faunal movement. As different plant species attract different fauna, the plant composition of the area may explain the differences in spore numbers at the various sampling locations. Canopy structure could have also affected the number of spores at all the locations sampled. The vegetative stands, composed of different species, formed a dense canopy cover that may have filtered the airborne spores and reduced the quantity of spores settling on the soil. This assertion though plausible, is not conclusive as canopy-level spore numbers were not assessed. The differences in the spore numbers across the sampling periods for the control locations did not differ from each other which may be due to the fact that with the exception of the canopy cover and faunal transport, no other factor which impact spore numbers acted significantly over the period. As no tillage occurred within the control areas, the changes in spore numbers were marginal, with relatively stable loads of spores. For the TSF however, cultural practices especially tillage is a principal factor affecting spore numbers. Generally, it can be posited that for the TSF, canopy cover, tillage and especially plant type and phenology influence the spore numbers. This agrees with Asmah’s (1996) assertion that the presence of mycotrophic plants, such as oil palm and Vertiver grass could possibly be the reason for the high spore numbers per gram of soil. The statistically significant variations in the number of spores over the sampled period for both tailing and control samples in this study support the observation of An et al. (2008) that seasonality has an effect on the number of AMF spores. No significant differences were found in the spore densities of AMF between the control site, which is a forest with minimal anthropogenic impact, and the TSF which undergoes periodic tillage. This points to the fact that fungal spores are ubiquitous in nature.
It is evident in the present study that diverse species of AMF were present in both the tailing and control samples, with the isolated spore types indicating that the fungi present consisted of different species of the known genera of mycorrhizal fungi at different stages of maturation. This observation is consistent with that of Asmah (1996). The AMF diversity did not depart from that obtained by Chaurasia et al. (2005) and Chaurasia and Khare (2006) who reported 16 and 15 AMF species respectively. The most abundant genus found in both the tailing and control samples was Glomus, which made up 33.2% of the specimens identified, with an occurrence of 100%. This was followed by Acaulospora, Gigaspora, Rhizophagus and Septoglomus. The dominant occurrence of the Glomus species has been shown to be as a result of their remarkable adaptation to different and sometimes drastic conditions as reported by Soni and Vyas, (2011) and Jha et al. (2011) thus, making them ubiquitous.
Seven common species – A. tuberculata, C. luteum, G. decipiens, G. ambisporum, G. marcocarpum, R. aggregatus and S. constrictum – were found in both the tailing and control samples and were very ubiquitous in most of the samples. This finding is consistent with that of Öpik et al., (2006) who observed that some AMF species such as G. mosseae, G. intraradices and G. etunicatum, are very frequent in their occurrence and have global distribution and thus highly ubiquitous, which were classified as “generalists” (Oehl et al., 2003). These seven species were found in areas that were several kilometers apart, with no direct linkage and very different soil characteristics. Thus, no matter the substrate or environmental conditions, these species will occur in the soil. On the other hand, D. nigra was found only in the control samples whilst nine other species – A, denticulate, A. foveate, D. epigea, G. cerebriformes, R. gregaria, R. aggregatus, R. clarus, S. deserticola and S. calospora – were found in only the tailing samples. These species are referred to as ‘Specialists’ in the sense that they occur in specific ecosystems or growth substrates such as the TSF and under specific nutrient, acid or neutral to alkaline soil conditions (Sieverding, 1991; Oehl et al., 2003). The presence of the generalist and specialist AMF observed in this study is in line with the observations of Mäder et. al. (2002), Jansa et. al. (2003) and Ambili et. al. (2012). Oehl et. al. (2003) are of the view that ‘generalists’ have been able to invade most existing ecosystems and establish a permanent presence whereas the specialists need specific ecological conditions to survive. Previous studies have also found members of the Gigasporaceae family, which includes Scutellospora, to be more prevalent in sandy soils (Cuenca & Lovera, 2010; Chaudhary et. al., 2014).
The AMF diversity at both sites and the various sampling locations can be attributed to localised different soil/substrate properties (Karaarslan & Uyanoz, 2011), type of vegetation (Hausmann & Hawkes, 2010) and most especially, soil moisture content (Shukla et. al., 2012). However, soil moisture content did not seem to have a huge impact in this study. The control soil was gravelly, more aerated, at the summit of a ridge and very dry as a result of low water holding capability arising from the gravelly nature of the soil and low levels of precipitation during the study. This, according to Shukla et. al., (2012) should provide enough space for AMF sporulation. On the other hand, the crushing of the gold-bearing ore in the beneficiation process results in the production of very fine particles which compacts after settling in the TSF. Thus, tailing exhibit the characteristics of a mixture of clay and loam. With these characteristics, dry tailing has the capacity to retain water, thus moisture remains in the tailing material for a long time and may not provide enough space for AMF proliferation as deduced from Dubey et. al., (2008). It was thus expected that the differences in the AMF diversity between the tailing and control samples, should have been statistically significant. As this was not observed in the present study, it can be suggested that moisture did not have a huge effect on the diversity.
The results of the study indicate that the spores isolated are heterogeneous in age and taxa, which is in line with the observation of An et. al. (2008) that spore types in field soils are heterogeneous in age, taxa and viability; with viability been seen as a potential indicator of changes in mycorrhizal activity. The heterogeneity in age is seen in the mix of young and senesce spores observed in all samples analysed whilst the different species and genera isolated give credence to the taxonomic heterogeneity. It can be said that the tailing samples do not depart significantly in terms of spore diversity from field soils.
The high mean Simpson’s Index of diversity (0.86±0.05) for the tailing samples shows shared dominance of many AMF species while the relatively lower value (0.62±0.08) indicated dominance of few species. This compares favourably with that of the control locations (0.81±0.05 and 0.64±0.02). Sixty out of the tailing sampling locations and 75% of the control locations recorded Simpson’s indices greater than 0.70, an indication of high diversity for this study. This observation agrees with those of Ambili et al. (2012) and Rajeshkumar et al. (2015). The Shannon-Weiner indices recorded for this study were much lower than those (1.91 and 4.56) recorded by both Ambili et al. (2012) and Rajeshkumar et al. (2015) respectively. This is so as species dominance in this study was shared by maximum of 10 out of the total 17 AMF species in the tailing samples whereas 5 out of 8 AMF shared the dominance in the control samples. The statistically significant differences in all the measured indices at the various sampling locations indicate that different practices and factors may be responsible for the distribution of AMF and that localized factors that affect the AMF community exist.
Abbott and Gazey (1994) suggest that the intensity of soil use modifies AMF species composition hence affecting diversity whilst Jansa et al. (2003) contend that factors such as the disturbance of AM fungal hyphal networks, changes in the nutrient content or weed populations (vegetation effects) are the driving forces affecting the AMF community structure. For this study, it can be posited that soil/substrate properties (Xu et al., 2018), plant species (Straker et al., 2007), dispersal mechanism and the means of spore propagation (Mangan & Adler, 2002) are critical to the incidence and abundance of AMF spores in the TSF.
CONCLUSION
This study revealed a high AMF diversity in the TSF. The communities of AMF inhabiting the tailings and natural soil were spatially heterogenous, indicating spatial variations. The variability patterns between the AMF in the tailing and natural soils are similar, an indication that AMF spores are ubiquitous. The mine tailings were inhabited by both generalist and specialist AMF. Seven common species (generalists) were found in both the tailing and control samples while nine other species were found only in the tailing samples thus making them “specialists”. The most abundant genus found in both the tailing and control samples was Glomus. These findings are important pointers for the adoption of mycorrhization in vegetation establishment and ultimately rehabilitation of decommissioned tailings storage facilities in Ghana. Further studies should however be carried out to determine how the various vegetation types found on the TSF affect AMF diversity and density.
REFERENCES
- Abbott, L. K., & Gazey, C. (1994). An ecological view of the formation of VA mycorrhizas. Plant and Soil, 159(1), 69. https://doi.org/10.1007/BF00000096
- Aghetara, J. B. (2012). Effects of artisanal and small scale mining on the environment and livelihoods in Ghana: a case study of selected mining communities in the Upper East Region [Thesis]. http://ir.knust.edu.gh:8080/handle/123456789/7484
- Akabzaa, T. (2009). Mining in Ghana: implications for national economic development and poverty reduction. Mining in Africa: Regulation and Development, 25–65.
- Ambili, K., Thomas, G. V., Indu, P., Gopal, M., & Gupta, A. (2012). Distribution of Arbuscular Mycorrhizae Associated with Coconut and Arecanut Based Cropping Systems. Agricultural Research, 1(4), 338–345. https://doi.org/10.1007/s40003-012-0036-4
- An, G.-H., Miyakawa, S., Kawahara, A., Osaki, M., & Ezawa, T. (2008). Community structure of arbuscular mycorrhizal fungi associated with pioneer grass species Miscanthus sinensis in acid sulfate soils: Habitat segregation along pH gradients. Soil Science and Plant Nutrition, 54(4), 517–528. https://doi.org/10.1111/j.1747-0765.2008.00267.x
- Ardestani, M. M., Jílková, V., Bonkowski, M., & Frouz, J. (2019). The effect of arbuscular mycorrhizal fungi Rhizophagus intraradices and soil microbial community on a model plant community in a post-mining soil. Plant Ecology, 220(9), 789–800. https://doi.org/10.1007/s11258-019-00953-w
- Asmah, A. E. (1996). Preliminary studies on the occurrence and infectivity of vesicular-arbuscular mycorrhizal fungi in some Ghanaian soils. Ghana Journal of Science, 36(1).
- Baah, A. (2005). Assessing Labour and Environmental Standards in South African Multinational Companies in the Mining Industry in Africa: A case of Gold Fields Ghana. Mining Africa: South African MNCs Labour and Social Performance. Namibia: National Labour and Economic Development Institute on Behalf of African Labour Research Network.
- Bever, J. D., Schultz, P. A., Pringle, A., & Morton, J. B. (2001). Arbuscular Mycorrhizal Fungi: More Diverse than Meets the Eye, and the Ecological Tale of WhyThe high diversity of ecologically distinct species of arbuscular mycorrhizal fungi within a single community has broad implications for plant ecology. BioScience, 51(11), 923–931. https://doi.org/10.1641/0006-3568(2001)051[0923:AMFMDT]2.0.CO;2
- Boulet, M. P., & Larocque, A. C. L. (1998). A comparative mineralogical and geochemical study of sulfide mine tailings at two sites in New Mexico, USA. Environmental Geology,33(2–3), 130– 142. https://doi.org/10.1007/s002540050233
- Carrasco, L., Gattinger, A., Fließbach, A., Roldán, A., Schloter, M., & Caravaca, F. (2010). Estimation by PLFA of Microbial Community Structure Associated with the Rhizosphere of Lygeum spartum and Piptatherum miliaceum Growing in Semiarid Mine Tailings. Microbial Ecology, 60(2), 265–271. JSTOR. https://doi.org/10.1007/s00248-009-9619-4
- Chaudhary, V. B., O’Dell, T. E., Rillig, M. C., & Johnson, N. C. (2014). Multiscale patterns of arbuscular mycorrhizal fungal abundance and diversity in semiarid shrublands. Fungal Ecology, 12, 32–43. https://doi.org/10.1016/j.funeco.2014.06.003
- Chaurasia, B., & Khare, P. K. (2006). Hordeum vulgare: a suitable host for mass production of arbuscular mycorrhizal fungi from natural soil. Applied Ecology and Environmental Research, 4(1), 45–53.
- Chaurasia, B., Pandey, A., & Palni, L. M. S. (2005). Distribution, colonization and diversity of arbuscular mycorrhizal fungi associated with central Himalayan rhododendrons. Forest Ecology and Management, 207(3), 315–324. https://doi.org/10.1016/j.foreco.2004.10.014
- Cicatelli, A., Torrigiani, P., Todeschini, V., Biondi, S., Castiglione, S. & Lingua, G. (2014). Arbuscular mycorrhizal fungi as a tool to ameliorate the phytoremediation potential of poplar: biochemical and molecular aspects. iForest 7: 333-341. – doi: 10.3832/ifor1045-007
- Cuenca, G., & Lovera, M. (2010). Seasonal variation and distribution at different soil depths of arbuscular mycorrhizal fungi spores in a tropical sclerophyllous shrubland. Botany, 88(1), 54–64. https://doi.org/10.1139/B09-100
- Dalpé, Y., & Aiken, S. G. (1998). Arbuscular mycorrhizal fungi associated with Festuca species in the Canadian High Arctic. Canadian Journal of Botany, 76(11), 1930–1938. https://doi.org/10.1139/b98-165
- Davison, J., Öpik, M., Daniell, T. J., Moora, M., & Zobel, M. (2011). Arbuscular mycorrhizal fungal communities in plant roots are not random assemblages: Selectivity in AMF-plant associations. FEMS Microbiology Ecology, 78(1), 103–115. https://doi.org/10.1111/j.1574-6941.2011.01103.x
- De La Cruz, R. E. (1991). Final report of the consultant on mycorrhiza program development in the IUC Biotechnology Center. PAU-IPB, Bogor, 168.
- Dubey, A., Mishra, M. K., Singh, P. K., & Vyas, D. (2008). Occurrence of AM fungi at varying stages of growth of rice plants. PROCEEDINGS OF THE NATIONAL ACADEMY OF SCIENCES INDIA SECTION B-BIOLOGICAL SCIENCES, 78, 51–55.
- Essah, M. (2021). Gold mining in Ghana and the UN Sustainable Development Goals: Exploring community perspectives on social and environmental injustices. Sustainable Development, n/a(n/a). https://doi.org/10.1002/sd.2233
- Gaur, A., & Adholeya, A. (2004). Prospects of arbuscular mycorrhizal fungi in phytoremediation of heavy metal contaminated soils. Current Science, 528–534.
- Gerdemann, J. W., & Nicolson, T. H. (1963). Spores of mycorrhizal Endogone species extracted from soil by wet sieving and decanting. Transactions of the British Mycological Society, 46(2), 235–244. https://doi.org/10.1016/S0007-1536(63)80079-0
- González, R. C., & González-Chávez, M. C. A. (2006). Metal accumulation in wild plants surrounding mining wastes. Environmental Pollution, 144(1), 84–92. https://doi.org/10.1016/j.envpol.2006.01.006
- Hausmann, N. T., & Hawkes, C. V. (2010). Order of plant host establishment alters the composition of arbuscular mycorrhizal communities. Ecology, 91(8), 2333–2343. https://doi.org/10.1890/09-0924.1
- (2015). Land acquisition and resettlement: Lessons learned. https://www.icmm.com/document/9714
- INVAM, W. (2014). International Culture Collection of (Vesicular) Arbuscular Mycorrhizal Fungi. Morgantown, WV: INVAM.
- Jansa, J., Mozafar, A., Kuhn, G., Anken, T., Ruh, R., Sanders, I. R., & Frossard, E. (2003). Soil tillage affects the community structure of mycorrhizal fungi in maize roots. Ecological Applications, 13(4), 1164–1176.
- Jha, A., Kumar, A., Kamalvanshi, M., & Shukla, A. (2011). Occurrence of arbuscular mycorrhizal fungi in rhizosphere of selected agroforestry tree species of Bundelkhand region. Indian Phytopathology, 64(2), 186.
- Johnson, N. C. (2010). Resource stoichiometry elucidates the structure and function of arbuscular mycorrhizas across scales: Tansley review. New Phytologist, 185(3), 631–647. https://doi.org/10.1111/j.1469-8137.2009.03110.x
- Johnson, N. C., Angelard, C., Sanders, I. R., & Kiers, E. T. (2013). Predicting community and ecosystem outcomes of mycorrhizal responses to global change. Ecology Letters, 16, 140–153.
- Karaarslan, E., & Uyanoz, R. (2011). Occurrence of arbuscular mycorrhizal fungi in some native plants grown on saline soils around the lake Tuz in Turkey and its relations with some physical and chemical properties of soil. Scientific Research and Essays, 6(20), 4238–4245.
- Kemp, D., Owen, J. R., & Lèbre, É. (2021). Tailings facility failures in the global mining industry: Will a ‘transparency turn’ drive change? Business Strategy and the Environment, 30(1), 122–134. https://doi.org/10.1002/bse.2613
- Klironomos, J., Zobel, M., Tibbett, M., Stock, W. D., Rillig, M. C., Parrent, J. L., Moora, M., Koch, A. M., Facelli, J. M., Facelli, E., Dickie, I. A., & Bever, J. D. (2011). Forces that structure plant communities: quantifying the importance of the mycorrhizal symbiosis. The New Phytologist, 189(2), 366–370.
- Krzaklewski, W., & Pietrzykowski, M. (2002). Selected Physicochemical Properties of Zinc and Lead Ore Tailings and their Biological Stabilisation. Water, Air, and Soil Pollution, 141(1), 125–141. https://doi.org/10.1023/A:1021302725532
- Kumar, P., Dobriyal, M., Kale, A., Pandey, A. K., Tomar, R. S., Thounaojam, E. (2022). Calculating forest species diversity with information-theory based indices using sentinel-2A sensors of Mahavir Swami Wildlife Sanctuary. PLoS ONE 17(5): e0268018. https://doi.org/10.1371/journal.pone.0268018
- Kumar, S., & Singh, J. (2019). Impact of Arbuscular Mycorrhizal Fungi (AMF) in Global Sustainable Environments. In A. N. Yadav, S. Singh, S. Mishra, & A. Gupta (Eds.), Recent Advancement in White Biotechnology Through Fungi (pp. 419–436). Springer International Publishing. https://doi.org/10.1007/978-3-030-25506-0_17
- Lekberg, Y., & Koide, R. T. (2005). Is plant performance limited by abundance of arbuscular mycorrhizal fungi? A meta-analysis of studies published between 1988 and 2003. New Phytologist, 168(1), 189–204. https://doi.org/10.1111/j.1469-8137.2005.01490.x
- Lottermoser, B. (2007). Mine Wastes. Springer-Verlag Berlin Heidelberg.
- Mäder, P., Fliessbach, A., Dubois, D., Gunst, L., Fried, P., & Niggli, U. (2002). Soil fertility and biodiversity in organic farming. Science, 296(5573), 1694–1697.
- Mangan, S. A., & Adler, G. H. (2002). Seasonal dispersal of arbuscular mycorrhizal fungi by spiny rats in a neotropical forest. Oecologia, 131(4), 587–597. https://doi.org/10.1007/s00442-002-0907-7
- McQuilken, J., & Hilson, G. (2016). Artisanal and small-scale gold mining in Ghana. Background Research to Inform an Action Dialogue. IIED, London.
- Mendez, M. O., Glenn, E. P., & Maier, R. M. (2007). Phytostabilization potential of quailbush for mine tailings. Journal of Environmental Quality, 36(1), 245–253.
- Mendez, M. O., & Maier, R. M. (2008). Phytoremediation of mine tailings in temperate and arid environments. Reviews in Environmental Science and Bio/Technology, 7(1), 47–59. https://doi.org/10.1007/s11157-007-9125-4
- Oehl, F., Sieverding, E., Ineichen, K., Mader, P., Boller, T., & Wiemken, A. (2003). Impact of Land Use Intensity on the Species Diversity of Arbuscular Mycorrhizal Fungi in Agroecosystems of Central Europe. Applied and Environmental Microbiology, 69(5), 2816–2824. https://doi.org/10.1128/AEM.69.5.2816-2824.2003
- Ofosu, G., Dittmann, A., Sarpong, D., and Botchie, D (2020). Socio-economic and environmental implications of Artisanal and Small-scale Mining (ASM) on agriculture and livelihoods. Sci. Policy106, 210–220. doi: 10.1016/j.envsci.2020.02.005
- Öpik, M., Moora, M., Liira, J., & Zobel, M. (2006). Composition of root-colonizing arbuscular mycorrhizal fungal communities in different ecosystems around the globe. Journal of Ecology, 94(4), 778–790. https://doi.org/10.1111/j.1365-2745.2006.01136.x
- Prayudyaningsih, R., Sari, R., & Mangopang, A. D. (2019). Isolation of Iindigenous arbuscular mycorrhizal fungi (AMF) to support revegetation on the nickel post-mining land. IOP Conference Series: Earth and Environmental Science, 308, 012038. https://doi.org/10.1088/1755-1315/308/1/012038
- Rajeshkumar, P. P., Thomas, G. V., Gupta, A., & Gopal, M. (2015). Diversity, richness and degree of colonization of arbuscular mycorrhizal fungi in coconut cultivated along with intercrops in high productive zone of Kerala, India. Symbiosis, 65(3), 125–141. https://doi.org/10.1007/s13199-015-0326-2
- Ros–Tonen, M. A. F., Aggrey, J. J., Somuah, D. P., & Derkyi, M. (2021). Human insecurities in gold mining: A systematic review of evidence from Ghana. The Extractive Industries and Society, 8(4), 100951. https://doi.org/10.1016/j.exis.2021.100951
- Schenck, N. C., & Perez, Y. (1990). Manual for the identification of vesicular mycorrhizal fungi. Florida, Synergistic Publications, Gainesville.
- Sharma, A., & Sharma, H. (2013). Role of vesicular arbuscular mycorrhiza in the mycoremediation of heavy toxic metals from soil. International Journal of Life Sciences Biotechnology and Pharma Research, 2, 2418–2431.
- Shukla, A., Kumar, A., Jha, A., Dhyani, S. K., & Vyas, D. (2012). Cumulative effects of tree-based intercropping on arbuscular mycorrhizal fungi. Biology and Fertility of Soils, 48(8), 899–909. https://doi.org/10.1007/s00374-012-0682-5
- Sieverding, E. (1991). Vesicular arbuscular mycorrhiza in tropical agrosystem. Deutsche Gesellsschaft Fur Techniische Zusammenarbeit GTZ) GMBH, Federal Republic of Germany.
- Silva, D.F., Moreira, J.V., Sousa, L.I.S., Santana, M.C., Mota, J.C.A., Queiroz, A.S., Nascimento, I.V., Silva, A.M.M., Araújo, A.S.F., Melo, V.M.M., Medeiros, E.V., Cardoso, E.J.B.N. & Pereira, A.P.A (2022). Arbuscular mycorrhizal fungi community in soils under desertification and restoration in the Brazilian semiarid. Res., 264.
- Silva-Flores, P., Bueno, C. G., Neira, J., & Palfner, G. (2019). Factors Affecting Arbuscular Mycorrhizal Fungi Spore Density in the Chilean Mediterranean-Type Ecosystem. Journal of Soil Science and Plant Nutrition, 19(1), 42–50. https://doi.org/10.1007/s42729-018-0004-6
- Smith, S. E., & Read, D. J. (2010). Mycorrhizal Symbiosis. Academic Press.
- Soni, P. K., & Vyas, D. (2011). Arbuscular mycorrhizal association with important medicinal plants of Sagar+. Indian Phytopathology.
- Spellerberg, I. F., & Fedor, P. J. (2003). A tribute to Claude Shannon (1916–2001) and a plea for more rigorous use of species richness, species diversity and the ‘Shannon–Wiener’Index. Global Ecology and Biogeography, 12(3), 177–179.
- Straker, C. J., Weiersbye, I. M., & Witkowski, E. T. F. (2007). Arbuscular mycorrhiza status of gold and uranium tailings and surrounding soils of South Africa’s deep level gold mines: I. Root colonization and spore levels. South African Journal of Botany, 73(2), 218–225. https://doi.org/10.1016/j.sajb.2006.12.006
- Titshall, L. W., Hughes, J. C. & Bester, H. C. (2013). Characterisation of alkaline tailings from a lead/zinc mine in South Africa and evaluation of their revegetation potential using five indigenous grass species. S Afr J Plant Soil 30:97–105
- Wang, Y., Narayanan, M., Shi, X., Chen, X., Li, Z., Natarajan, D. & Ma, Y. (2022). Plant growth-promoting bacteria in metal-contaminated soil: Current perspectives on remediation mechanisms. Frontiers in Microbiology,13
- Xu, J., Liu, S., Song, S., Guo, H., Tang, J., Yong, J. W. H., Ma, Y., & Chen, X. (2018). Arbuscular mycorrhizal fungi influence decomposition and the associated soil microbial community under different soil phosphorus availability. Soil Biology and Biochemistry, 120, 181–190. https://doi.org/10.1016/j.soilbio.2018.02.010
- Yang, H., Zhang, Q., Koide, R. T., Hoeksema, J. D., Tang, J., Bian, X., Hu, S., & Chen, X. (2017). Taxonomic resolution is a determinant of biodiversity effects in arbuscular mycorrhizal fungal communities. Journal of Ecology, 105(1), 219–228. https://doi.org/10.1111/1365-2745.12655
- Yankson, P. W., & Gough, K. V. (2019). Gold in Ghana: The effects of changes in large-scale mining on artisanal and small-scale mining (ASM). The Extractive Industries and Society, 6(1), 120–128.
- Youden, W. J., & Mehlich, A. (1937). Selection of Elficient Methods for Soil Sampling. Contributions From.
- Yiridomoh, G. Y. (2021). “Illegal” Gold Mining Operations in Ghana: Implication for Climate- Smart Agriculture in Northwestern Ghana. Sustain. Food Syst. 5:745317. doi: 10.3389/fsufs.2021.745317
- Zhao, Z. W., Wang, G. H., & Yang, L. (2003). Biodiversity of arbuscular mycorrhizal fungi in a tropical rainforest of Xishuangbanna, southwest China. Fungal Divers, 13, 233–242.
- Zhao, Z.-W., Xia, Y.-M., Qin, X.-Z., Li, X.-W., Cheng, L.-Z., Sha, T., & Wang, G.-H. (2001). Arbuscular mycorrhizal status of plants and the spore density of arbuscular mycorrhizal fungi in the tropical rain forest of Xishuangbanna, southwest China. Mycorrhiza, 11(3), 159–162. https://doi.org/10.1007/s005720100117