
Genetic Profiling and Prevalence of Kelch 13 Propeller Gene Mutations in Plasmodium Falciparum Isolates from Nasarawa North Senatorial District, Nigeria
- Timshana Emmanuel
- Ngwai Yakubu Boyi
- Igbawua Isaac Nyiayem
- 635-646
- Apr 7, 2025
- Medical Research
Genetic Profiling and Prevalence of Kelch 13 Propeller Gene Mutations in Plasmodium Falciparum Isolates from Nasarawa North Senatorial District, Nigeria
Timshana Emmanuel*, Ngwai Yakubu Boyi, Igbawua Isaac Nyiayem
Department of Microbiology, Nasarawa State University, PMB 1022, Keffi, Nasarawa State
*Corresponding Author
DOI: https://doi.org/10.51244/IJRSI.2025.12030047
Received: 13 December 2024; Accepted: 04 January 2025; Published: 07 April 2025
ABSTRACT
Malaria continues to pose a significant public health challenge in Nasarawa North Senatorial District, Nigeria, with increasing concerns about drug resistance. This study investigated the prevalence and genetic diversity of Plasmodium falciparum by analyzing Kelch 13 propeller gene mutations, a key marker for artemisinin resistance. A cross-sectional study was conducted across selected healthcare facilities, where 384 venous blood samples were collected and analyzed using microscopy, rapid diagnostic test kits (RDTs), and polymerase chain reaction (PCR). A total of 107 P. falciparum-positive cases were identified, and 27 samples were successfully sequenced. The results revealed a P. falciparum prevalence of 27.9%, with a mutation prevalence of 11.11%, with three-point mutations (D57V, A158C, and A157G) detected from P. falciparum isolates. General Hospital Akwanga recorded the highest mutation frequency (7.40%), while no Kelch 13 mutations were detected at General Hospital Nasarawa Eggon and Wamba. Statistical analysis (χ² = 8.02, p = 0.046) showed a significant variation in mutation distribution across healthcare facilities, pointing to area-specific factors influencing mutation prevalence. These findings emphasize the importance of continuous genetic surveillance to monitor resistance trends and guide malaria control strategies. Further research should explore the functional implications of Kelch 13 mutations to assess their potential impact on artemisinin efficacy.
Key words: Kelch 13; Malaria; Artemisinin; Nasarawa; Sequencing; RDT
INTRODUCTION
Malaria remains a major public health challenge, particularly in Africa, where the disease burden is highest. In 2018, there was an estimated 228 million malaria cases worldwide, with the African region accounting for 93% (213 million) of these cases. The Southeast Asia Region and the Eastern Mediterranean Region followed with 3.4% and 2.1%, respectively. Malaria-related deaths were estimated at 405,000 globally, with Plasmodium falciparum responsible for over 90% of fatalities [1]..
The African continent remains at the epicenter of the malaria crisis. In 2022, it accounted for 94% (233 million) of global malaria cases and 95% of malaria-related deaths, with young children under five years, being the most vulnerable, as they constituted 80% of malaria deaths in the region.. The devastating impact of malaria in Africa is further reflected in the estimated 608,000 malaria deaths worldwide in 2022, with the majority occurring in the African region. Nigeria, along with the Democratic Republic of the Congo, Niger, and Tanzania, accounted for nearly half of all malaria deaths globally in 2022 [2].
Effective antimalarial drugs play a crucial role in malaria control by ensuring rapid parasite clearance when infections are detected early [3]. Unfortunately, P. falciparum, the most virulent malaria parasite, has developed resistance to most antimalarial drugs [4]. Chloroquine (CQ), once the primary treatment for uncomplicated P. falciparum malaria, was replaced by Sulfadoxine-Pyrimethamine (SP) due to resistance. However, widespread resistance to SP by the mid-1990s rendered the drug ineffective, leading to the adoption of Artemisinin-based Combination Therapies (ACTs) as the preferred treatment [3].
To slow the development of drug resistance, the World Health Organization (WHO) recommended the use of ACTs in malaria-endemic countries. Nigeria introduced ACTs in 2005, designating artemether–lumefantrine (AL) as the first-line treatment for uncomplicated malaria, with Artesunate + Amodiaquine as an alternative. Artemisinin derivatives, including artesunate, artemether, and dihydroartemisinin (DHA), exhibit rapid parasite clearance and are well tolerated, especially when combined with long-acting partner drugs in ACT regimens [4].
Over the past two decades, there has been a significant decline in malaria cases in Africa, largely attributed to the widespread implementation of ACTs [5]. However, this progress is now under threat due to the emergence of ACT-resistant P. falciparum strains. Resistance, initially detected in the Greater Mekong Subregion, has now spread globally, undermining malaria control efforts [6]. Specific mutations in the P. falciparum Kelch 13 (pfk13) propeller domain, including N458Y, Y493H, R539T, I543T, and C580Y, have been associated with delayed parasite clearance following artemisinin treatment, particularly in Southeast Asia [7].
The identification of pfk13 mutations linked to artemisinin resistance highlights the importance of molecular surveillance in malaria-endemic regions [8]. While Kelch 13 mutations are widespread in Southeast Asia, studies indicate that mutations in Africa are distinct, with weaker selective pressure observed. The role of antimalarial drug use in shaping these mutation patterns in African P. falciparum strains remains uncertain. Several studies have reported multiple Kelch 13 mutations across Africa, including non-synonymous mutations (M472I, Y558C, K563R, P570L, and P615S) in Niger, a novel mutation (R622I) in Ethiopia, and 15 non-synonymous mutations in Senegal. The P553L mutation has been identified in Mali, Kenya, and Malawi [7]. However, these mutations have not been linked to delayed parasite clearance [7].
Despite evidence of Kelch 13 polymorphisms in various countries, data on the presence and impact of these mutations in Nigeria remain scarce [5]. This informs the need for continuous molecular surveillance to monitor the evolution of Kelch 13 mutations and their potential implications for ACT efficacy.
MATERIALS AND METHODS
Study area
Nasarawa North Senatorial District in Nasarawa State, Nigeria, comprises three Local Government Areas (LGAs): Akwanga, Nasarawa Eggon, and Wamba, with a combined population of approximately 335,453 residents. Nasarawa State lies between latitudes 7°45’–9°25′ N and longitudes 7°–9°37′ E, sharing boundaries with Kaduna, Plateau, Taraba, Benue, Kogi States, and the Federal Capital Territory (FCT) [9]. The study was cross-sectional and health facility-based, conducted in General Hospital Akwanga, General Hospital. Nasarawa Eggon, OIA Hospital Akwanga, and General Hospital Wamba. The region is malaria-endemic, with seasonal transmission that peaks during the rainy season. Its diverse topography, consisting of hills, undulating plains, and lowlands, makes it a critical area for malaria research and control strategies.
Study Population
Nasarawa North Senatorial District, located in Nasarawa State, Nigeria, comprises three Local Government Areas (LGAs): Akwanga, Nasarawa Eggon, and Wamba. The district has a combined population of approximately 335,453 residents. Nasarawa North Senatorial district has a total population of 335.453 which population distribution by Local Government Areas: Akwanga, 113,430; Nasarawa Eggon, 149,129, Wamba, 72,894.
Sample Size Determination
The sample size was determined using standard epidemiological formula as described by [10].:
N= Z2⋅p⋅ (1−p)
E2
Where: N= required sample size
Z = z-value (standard score) corresponding to the desired confidence level (e.g., 1.96 for 95% confidence)
p = estimated proportion of the population with the characteristic (0.5 was used to maximize the sample size)
E = margin of error (e.g., 0.05 for ±5%)
Given that: Z= 1.96, P= 0.5, E= 0.05
Sample size for this study was calculated as follows;
n= 1.96 x 1.96 x 0.5 (1 – 0.5) ÷ 0.05 x 0.05
n = 3.8416 x 0.5 x 0.5 ÷ 0.0025
n = 0.9604 ÷ 0.0025
n = 384.2
n = ~ 384 samples
In view of the above, a total of 384 venous blood samples were collected in EDTA bottles as follows; General Hospital Akwanga (GHA) = 96, OLA Hospital Akwanga (OLAA) = 96, General Hospital Nasarawa Eggon = 96, General Hospital Wamba = 96.
Ethical Approval
Ethical approval was granted by the Nasarawa State Ministry of Health Research Ethics Committee, ensuring compliance with approved ethical protocols. Additionally, written informed consent was obtained from all participants before their enrollment in the study.
Subject Selection
Inclusion Criteria
Only patients residing in Nasarawa North Senatorial District who presented with a body temperature of ≥37.5°C or a history of fever within the 24 hours prior to their visit were included in the study, provided they had not taken any antimalarial medication in the preceding week. Blood samples were deemed suitable for analysis if microscopic examination of Giemsa-stained thick films and Rapid Diagnostic Tests (RDTs) confirmed mono-infection with P. falciparum.
Exclusion Criteria
Patients were excluded from the study if their Giemsa-stained blood films showed no malaria parasites or if they had mixed infections. Additionally, individuals residing outside Nasarawa North Senatorial District were not included in the study.
Blood Collection
Venous blood samples were collected in EDTA tubes from study participants and prepared for microscopic examination to detect Plasmodium falciparum parasites. A total of 96 samples were collected from each health facility using standard protocols to ensure accuracy and reliability. The blood samples collected in EDTA from the study subjects were prepared and microscopically examined to detect malaria parasites (P. falciparum)
Laboratory Investigation
Microscopic Identification Malaria Parasites
A clean, grease-free glass slide was used to prepare thick and thin blood films from well-mixed whole blood collected in EDTA tubes. Two drops of blood were placed at one end of the slide. One drop was spread in a circular motion to create a thick film, while the second drop was used to make a thin film. A spreader slide was positioned at a 45° angle, brought into contact with the blood drop, and smoothly pushed forward to create an evenly distributed thin film. The prepared slide was labeled and air-dried. Thin films were then fixed with ethanol, allowing the solvent to evaporate before staining. The dried thick and thin films were stained using a freshly prepared 1:10 dilution of Giemsa stain in pH 7.2 buffer for 10 minutes. After staining and air drying, the films were examined under a light microscope using a 100x oil immersion objective as described [11]. Thin films were utilized for the identification of Plasmodium species and parasite developmental stages, while thick films were used to determine parasite density.
Rapid Diagnostic Testing for P. falciparum
Positive blood sample from microscopy were further screened for mono-infection with Plasmodium falciparum using a Rapid Diagnostic Test (RDT) specific for P. falciparum Histidine-Rich Protein 2 (HRP2) (Standard Diagnostics, Inc., Korea). To perform the test, 5µl of whole blood was dispensed into the sample well of a labeled, pre-warmed SD Bioline P. falciparum malaria antigen test device placed on a flat surface. Subsequently, two drops (100µl) of assay diluent were added to the diluent well, and the reaction was timed for 15 minutes. The results were interpreted thus: Positive samples displayed two distinct color bands—one in the control region and the other in the test line region; Negative samples exhibited a single color band in the control region only results were considered invalid if no color bands appeared in either region, or if a band appeared only in the test region without a corresponding control band.
Confirmed mono-infected samples were then spotted onto Whatman No. 3 filter paper (Whatman International Ltd., Maidstone, England), carefully labeled, and air-dried to form Dried Blood Spots (DBS). Each DBS sample was individually placed in a zip-lock bag with a desiccant to prevent moisture contamination and stored at ambient temperature. These samples were subsequently transported to Nucleometrix Research Laboratory, Yenagoa, Bayelsa State, for molecular analysis
Molecular Detection of P. falciparum Kelch 13 Gene
DNA extraction and Quantification
The parasite DNA was extracted from dried blood spot (DBS) samples using the Quick-DNA Miniprep Plus Kit (Zymo Research, USA) following the manufacturer’s protocol. Blood-impregnated Whatman filter paper (5 mm diameter) was perforated using a single-punch perforator and placed into ZR Bashing Bead™ Lysis tubes containing 400 µl of Bashing Bead Buffer (BBB). The samples were processed in a disruptor (Disruptor Genie) at maximum speed for 5 minutes, followed by centrifugation at 10,000 rpm for 1 minute.
To the lysate, 40 µl of Proteinase K enzyme and 360 µl of Solid Tissue Buffer (Blue) were added to degrade proteins, then incubated at 55°C for 15 minutes. After another centrifugation step (10,000 rpm for 1 min), 400 µl of the supernatant was mixed with 800 µl of Genomic Binding Buffer, vortexed, and transferred to Zymo-spin (IIC-XLR) columns for DNA purification via multiple wash steps using Pre-Wash Buffer (400 µl), g-DNA Wash Buffer (700 µl), and Wash Buffer (200 µl). Each step was followed by centrifugation at 12,000 rpm for 1 minute.
The spin columns were transferred to labeled microcentrifuge tubes, and 50 µl of DNA Elution Buffer was added. After a 5-minute incubation at room temperature, the tubes were centrifuged at 14,500 rpm for 1 minute, yielding purified DNA, which was stored for PCR amplification.
Finally, the extracted genomic DNA was quantified using the Nanodrop 1000 spectrophotometer, which was initialized with sterile distilled water and blanked using normal saline. Two microliters of extracted DNA were placed on the lower pedestal, the upper pedestal was lowered for contact, and DNA concentration was measured by clicking the “measure” button in the software.
Kelch 13 Gene PCR Amplification
The Kelch 13 gene of Plasmodium falciparum was amplified using Kelch 13-specific primers on an ABI 9700 Applied Biosystems thermal cycler in a 50 µL reaction volume for 35 cycles. The PCR reaction mixture consisted of 2X DreamTaq Master Mix (containing Taq polymerase, dNTPs, and MgCl₂) from Inqaba, South Africa, the primers at a concentration of 0.4M and the extracted genomic DNA as the template. The PCR cycling conditions were as follows: Initial denaturation: 95°C for 5 minutes; Denaturation: 95°C for 30 seconds; Annealing: 53°C for 30 seconds; Extension: 72°C for 30 seconds (for 35 cycles) and final extension: 72°C for 5 minutes.
Following amplification, the PCR products were electrophoresed on a 1.5% agarose gel at 120V for 15 minutes and visualized under a UV transilluminator to confirm successful amplification.
Table 1: Primer sequences and cycling conditions used to amplify K13-propeller gene of Plasmodium falciparum isolates and Nested PCR of Kelch 13 genes
| Target genes | Primer sequence (5‘–3′) | PCR cycling conditions | Amplicon size |
| Kelch 13 (P) | F: CGGAGTGACCAAATCTGGGA
R: GGGAATCTGGTGGTAACAGC |
94 °C 5 min/[94 °C 90 s, 55 °C 45 s, 72 °C , 90 s] × 40 cycles, 72 °C 10 min. | 2096bp
|
| Kelch 13 (S) | F: GCCAAGCTGCCATTCATTTG
R: GCCTTGTTGAAAGAAGCAGA |
94 °C 5 min/[94 °C 90 s, 55 °C 45 s, 72 °C 90 s] × 40 cycles, 72 °C 10 min | 848bp |
P = Primary PCR reaction, S = Secondary PCR reaction, F = Forward primer, R = Reverse primer, bp = base pairs. Source: Huang et al., [12]
Sequencing of Kelch 13 genes
Out of the 107 P. falciparum positive samples, 27 were sequenced using the BigDye Terminator kit on a 3510 ABI sequencer by Inqaba Biotechnological, Pretoria South Africa. The sequencing was done at a final volume of 10ul, the components included 0.25 ul BigDye® terminator v1.1/v3.1, 2.25ul of 5 x BigDye sequencing buffer, 10uM Primer PCR primer, and 2-10ng PCR template per 100bp. The sequencing conditions were as follows 32 cycles of 96°C for 10s, 55°C for 5s and 60°C for 4min
Phylogenetic Analysis
Obtained sequences were edited using the bioinformatics algorithm Trace edit, similar sequences were downloaded from the National Center for Biotechnology Information (NCBI) data base using BLASTN. These sequences were aligned using ClustalX. The evolutionary history was inferred using the Neighbor-Joining method in MEGA 6.0 [13]. The bootstrap consensus tree inferred from 500 replicates [14] is taken to represent the evolutionary history of the taxa analysed. The evolutionary distances were computed using the Jukes-Cantor method [15].
Statistical Analysis
Data analysis was performed using SPSS version 25, employing both descriptive statistics (e.g., percentages and frequencies) and inferential statistics to interpret findings. Chi-square tests were conducted to evaluate differences in mutation prevalence across study sites, with p < 0.05 considered statistically significant
RESULTS AND DISCUSSION
Results
The overall prevalence of Plasmodium falciparum in Nasarawa North Senatorial District was 27.86%, with 107 out of 384 samples testing positive. A slight gender difference was observed, with 55 cases (51.4%) in females and 52 cases (48.6%) in males; however, this difference was not statistically significant.
Analysis of prevalence by education level showed that individuals with no formal education had the highest prevalence (35.00%), followed by those with primary education (28.89%), secondary education (26.92%), and tertiary education (24.04%). Comparing urban and rural residence, P. falciparum prevalence was higher in rural areas (30.00%) than in urban areas (24.31%). This disparity may be due to increased exposure to mosquito breeding sites, lower utilization of insecticide-treated nets (ITNs), and limited access to healthcare services in rural communities.
Notably, P. falciparum prevalence varied across healthcare facilities. Ola Hospital Akwanga (33.3%) recorded the highest prevalence, while General Hospital Akwanga (16.7%) had the lowest. This variation shows the significance of tailored malaria control programs, especially in high-burden areas.
Among the 107 P. falciparum-positive cases, 27 samples were sequenced for Kelch 13 mutations, with 3 (11.1%) testing positive. This indicates a relatively low prevalence of Kelch 13 mutations within the study population. Three specific point mutations detected were: D57V, A158C, and A157G. The distribution of these mutations varied across healthcare facilities, with General Hospital Akwanga (GHA) showing the highest prevalence at 7.40%, followed by OLA Hospital Akwanga (OLAA) at 3.70%. No Kelch 13 mutations were detected in General Hospital Nasarawa Eggon (GHN) and General Hospital Wamba (GHW). The variation in mutation distribution was statistically significant (χ² = 8.02, p = 0.046).
Although the identified Kelch 13 mutations (D57V, A158C, and A157G) are not among the validated WHO markers of artemisinin resistance (such as C580Y, Y493H, or R539T), their location within the Kelch 13 propeller domain suggests they could be pre-resistance markers or compensatory mutations, potentially stabilizing resistance-associated mutations. The prevalence of p. falciparum is shown in table 1 and 2, Prevalence of Kelch 13 Mutations across Health facilities fig 1 while plate 1, Agarose gel electrophoresis shows the amplified Kelch 13 and fig, 2 shows Sequence alignment of the protein sequence of the Kelch 13 gene showing 3 points mutations
Table 2: Prevalence of P. falciparum from study facilities in the study area
| Facilities | No. Exam -ined | No. Negative | No. of Positive males | No. of Positive females | Total No. Positive | % positive |
| GHA | 96 | 80 | 11 | 5 | 16 | 16.67 |
| OLAA | 96 | 64 | 13 | 19 | 32 | 33.33 |
| GHN | 96 | 67 | 17 | 12 | 29 | 30.21 |
| GHW | 96 | 66 | 11 | 19 | 30 | 31.25 |
| TOTAL | 384 | 277 | 52 | 55 | 107 | 27.86 |
GHA= General Hospital Akwanga, OLAA= Ola Hospital Akwanga, General Hospital Nasarawa Eggon, GHW= General Hospital Wamba
Table 3: Malaria Prevalence by Age Group, Education Level, and Residence
| Category | Subcategory | No. Tested | No. Negative | No. Positive | % Positive |
| Age Group (Years) | 0-5yrs | 65 | 38 | 27 | 41.54% |
| 6-14yrs | 90 | 64 | 26 | 28.89% | |
| 15-24yrs | 110 | 83 | 27 | 24.55% | |
| ≥25yrs | 119 | 92 | 27 | 22.69% | |
| Total | 384 | 277 | 107 | 27.86% | |
| Education Level | No Formal Education | 60 | 39 | 21 | 35.00% |
| Primary Education | 90 | 64 | 26 | 28.89% | |
| Secondary Education | 130 | 95 | 35 | 26.92% | |
| Tertiary Education | 104 | 79 | 25 | 24.04% | |
| Total | 384 | 277 | 107 | 27.86% | |
| Residence | Rural | 240 | 168 | 72 | 30.00% |
| Urban | 144 | 109 | 35 | 24.31% | |
| Total | 384 | 277 | 107 | 27.86% |
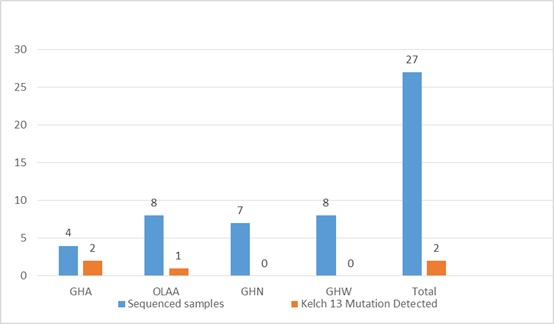
GHA= General Hospital Akwanga, OLAA= Ola Hospital Akwanga, General Hospital Nasarawa Eggon, GHW= General Hospital Wamba
Fig 1. Prevalence of Kelch 13 Mutations across Health facilities in Nasarawa North Senatorial District
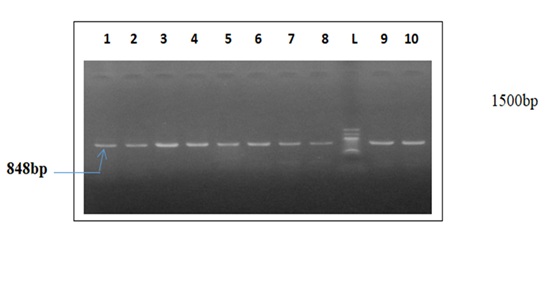
Figure 2: Agarose gel electrophoresis showing the amplified Kelch 13. Lanes 1- 8, 9 & 10 represent the Kelch 13 gene bands at 848bp while lane L represent the 100 bp DNA molecular ladder at 1500bp
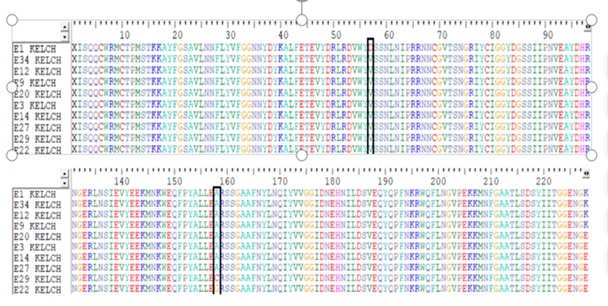
Fig 3: Sequence alignment of the protein sequence of the Kelch 13 gene showing 3 points mutations at positions D57V (sample E1), A158C (sample E29) and A157G (sample E22) (indicated by red colour in the masked black line).
DISCUSSION
Malaria remains a major health challenge in sub-Saharan Africa, with Plasmodium falciparum causing most infections and deaths [2]. Kelch13 gene mutations driving artemisinin resistance threaten treatment effectiveness [16]. In this study, the overall prevalence of P. falciparum in Nasarawa North Senatorial District was found to be 27.9% as shown in table 1. This is significantly higher than the 15.7% prevalence reported among pregnant women in Lafia, Nasarawa South [17]. These variations may be attributed to differences in, healthcare access, and socio-economic conditions across the areas.
Across Nigeria, malaria prevalence shows notable geographical disparities. For instance, Kano State reported a prevalence of 33.5%, with the highest infection rates observed among children under five years old, mirroring this study’s findings [18]. Conversely, Lagos State reported a prevalence of 22.7%, reflecting the urban advantage of improved healthcare infrastructure [5]. Beyond Nigeria, similar patterns emerge across Africa; the Democratic Republic of Congo documented a prevalence of 45.3% in rural communities, underscoring the heightened vulnerability of these populations due to limited access to preventive measures and medical intervention [19].
Age-stratified analysis revealed that children under five in Nasarawa North exhibited the highest infection rates (41.54%). This observation aligns with findings from Burkina Faso, where children aged 0-5 years showed a prevalence of 39.8%, attributed to their underdeveloped immunity [20]. Comparable trends have been reported in Tanzania, where younger children demonstrated increased susceptibility [21]. Moreover, disparities between rural and urban settings highlight the role of environmental and infrastructural factors in malaria transmission, as evidenced by the 30.0% prevalence in rural Nasarawa North compared to 24.3% in urban areas. Similar findings have been reported in Uganda, where rural regions experienced higher transmission rates [22].
Molecular analysis confirmed the presence of the kelch13 gene in P. falciparum isolates, with three mutations identified: D57V, A158C, and A157G. These findings parallel reports from Lagos, Nigeria, where kelch13 polymorphisms were observed, though at low frequencies [5]. In Ghana, the A578S mutation has been identified, raising analogous concerns about potential artemisinin resistance markers [23]. Furthermore, Rwanda has reported the R561H mutation, associated with delayed parasite clearance, indicating early signs of resistance [24]. Notably, Southeast Asia remains the epicenter of confirmed artemisinin-resistant kelch13 variants, such as C580Y, which has been strongly linked to treatment failures [6].
In other parts of the World, the increasing frequency of kelch13 mutations underscores the selective pressure exerted by artemisinin-based combination therapies (ACTs). In Cambodia, high mutation frequencies have been linked to delayed parasite clearance [25]. Even in places like the Amazon Basin, where artemisinin resistance has been slower to emerge, recent reports indicate the appearance of kelch13 mutations, pointing to the need for heightened global surveillance [26]. Across Africa, the spread of these mutations, though less prevalent than in Southeast Asia, demands proactive intervention [27].
Several studies have documented the role of kelch13 mutations as markers of artemisinin resistance. [28] reported a steady increase in mutation frequencies in Kenyan isolates, linked to regions with higher artemisinin drug pressure. Similarly, [22] found that kelch13 mutations in Uganda were more prevalent in areas with consistent ACT use, reflecting a direct correlation between drug pressure and mutation emergence. In contrast, areas with lower artemisinin use have shown fewer mutations, emphasizing the impact of drug selection pressure on parasite evolution.
These findings highlight the need for continuous molecular surveillance of kelch13 mutations to track resistance patterns across Africa. Studies from Senegal [29] reinforce the importance of integrating genetic monitoring into national malaria control programs to detect early signals of resistance and adapt treatment policies accordingly.
The findings from Nasarawa North shows the pressing need for sustained genetic surveillance of kelch13 mutations in Nigeria and the broader West African countries. This agrees with findings from Uganda and Kenya, where rising kelch13 mutations have been linked to prolonged artemisinin use [22] [28]. Reinforcing malaria control measures such as scaling up the distribution of insecticide-treated nets and bolstering healthcare infrastructure remains vital for curbing the spread of resistance.
Expanding evidence on the prevalence of Plasmodium falciparum and mutations in the kelch13 gene across Africa. Notably, mutations such as A675V and C469Y have been identified, with the prevalence of kelch13 mutations increasing from 3.9% in 2015 to 19.8% in 2019 [30]. Additionally, the A578S mutation, although not linked to artemisinin resistance, has been detected in multiple African countries [30]. These findings underscore the importance of ongoing surveillance to monitor the emergence and spread of these mutation
Table 2 revealed Age-stratified prevalence, with children under five having the highest infection rates (41.54%), consistent with findings from Kano State, where younger children showed increased susceptibility [16]. The disparity in infection rates between rural (30.0%) and urban (24.3%) populations in Table 2 indicates the influence of environmental and infrastructural factors in malaria transmission.
Plate 3 presents agarose gel electrophoresis results, illustrating the amplified kelch13 gene. Lanes 1-10 display kelch13 gene bands at 848bp, while lane L represents the 100bp DNA molecular ladder at 1500bp. The successful amplification of kelch13 confirms the presence of this gene in the analyzed P. falciparum isolates. Molecular analysis revealed three kelch13 mutations: D57V, A158C, and A157G. The presence of these mutations in P. falciparum isolates is consistent with findings from Lagos, Nigeria, where kelch13 polymorphisms were detected at varying frequencies [5]. While Southeast Asia remains the epicenter of confirmed artemisinin-resistant kelch13 variants [16], increasing reports of these mutations in African populations imply potential pre-resistance markers that warrant further surveillance [27].
A comparative assessment with global data highlights that kelch13 mutations are less frequent in West Africa than in Southeast Asia, where mutations such as C580Y have been strongly linked to delayed parasite clearance [6]. In Uganda, [22] observed an increasing prevalence of kelch13 mutations in response to sustained artemisinin drug pressure, raising concerns about future resistance patterns in Africa.
The findings from Nasarawa North emphasize the need for continued genetic surveillance of kelch13 mutations and P. falciparum prevalence across Nigeria and West Africa. Strengthening malaria control strategies, including increased use of insecticide-treated nets, improved access to healthcare, and monitoring of drug efficacy, will be crucial in mitigating emergence of resistance in malarial parasites.
Given the observed kelch13 mutations, further functional studies are needed to determine their impact on artemisinin sensitivity and treatment efficacy.
CONCLUSION
This study provides evidence of Kelch 13 mutations in P. falciparum isolates from Nasarawa North, with significant variation in mutation prevalence across health facilities. Although the identified mutations are not WHO-validated resistance markers, their presence warrants continuous monitoring to detect any emerging resistance trends. Strengthening malaria control programs and expanding genetic surveillance will be crucial for maintaining the efficacy of ACTs.
ACKNOWLEDGMENTS
The authors sincerely appreciate the Nasarawa State Ministry of Health, the participating healthcare facilities, and all study participants for their support. Special thanks to the Department of Microbiology, Nasarawa State University, Keffi, the Faculty of Natural and Applied Sciences, and the Postgraduate School for providing a conducive research environment. We also acknowledge the management of Nasarawa State College of Health Science and Technology, Keffi, and the technical support from Neuclomatrix Laboratory, Yenagoa. Finally, we are grateful to all who contributed in any capacity to the success of this research.
REFERENCES
- Dafalla OM., Hassan MM., & Mohamed SB. (2020). Global malaria burden and resistance patterns: An epidemiological overview. Tropical Medicine and Infectious Disease, 5(2), 89. https://doi.org/10.3390/tropicalmed5020089
- World Health Organization. (2023). World malaria report 2023. Retrieved from [https://www.who.int/publications/i/item/9789240076785]
- Ndwiga L., Wamae K., & Githinji G. (2021). Antimalarial resistance in sub-Saharan Africa: A review of emerging threats. Infectious Diseases and Therapy, 10(3), 1105-1122. https://doi.org/10.1007/s40121-021-00440-2
- World Health Organization. (2020). World malaria report 2020. Retrieved from [https://www.who.int/publications/i/item/9789240015791]
- Igbasi U, Okeke C, & Onyekachi E. (2019). Prevalence of Kelch13 mutations in falciparum isolates from Lagos, Nigeria. Malaria Journal, 18(1), 200.
- Ashley EA., Dhorda M, Fairhurst RM., Amaratunga C, Lim P, Suon S, … & Dondorp AM. (2014). Spread of artemisinin resistance in Plasmodium falciparum New England Journal of Medicine, 371(5), 411-423. https://doi.org/10.1056/NEJMoa1314981
- Abubakar U, Sulaiman SA, & Jimoh RO. (2020). Malaria chemotherapy and resistance in Africa: Challenges and prospects. Infectious Diseases of Poverty, 9(1), 27. https://doi.org/10.1186/s40249-020-00651-w
- Jeang L. (2024). Surveillance of antimalarial resistance markers in Africa. WHO Malaria Report, 2024. Retrieved from [https://www.who.int] (https://en.wikipedia.org/wiki/Nasarawa_State
- Naing L, Winn T, Rusli1 BN. Practical Issues in Calculating the Sample Size for prevalence studies. Archives of Orofacial Sciences. 2006; 1: 9 -14
- Cheesbrough M. (2006). District Laboratory Practice in Tropical Countries. (2nd edition update). Cambridge University Press : 248 – 249
- Huang B Deng C, Yang T, Xue L, Wang Q, Huang S, Su XZ, Liu Y, Zheng S, Guan Y, Xu Q, Zhou J, Yuan J, Bacar A, Abdallah KS, Attoumane R, Mliva AM, Zhong Y, Lu F, & Song J. Polymorphisms of the artemisinin resistant marker (K13) in Plasmodium falciparum parasite populations of Grande Comore Island 10 years after artemisinin combination therapy. Parasites & vectors. 2015; 8: 634. https://doi.org/10.1186/s13071-015-1253-z
- Saitou N, & Nei M. (1987). The neighbor-joining method: A new method for reconstructing phylogenetic trees. Molecular Biology and Evolution, 4(4), 406-425. https://doi.org/10.1093/oxfordjournals.molbev.a040454
- Felsenstein J. (1985). Confidence limits on phylogenies: An approach using the bootstrap. Evolution, 39(4), 783-791. https://doi.org/10.2307/2408678
- Jukes TH., & Cantor CR. (1969). Evolution of protein molecules. In H. N. Munro (Ed.), Mammalian Protein Metabolism (Vol. 3, pp. 21-132). Academic Press. https://doi.org/10.1016/B978-1-4832-3211-9.50009-7
- Ariey F, Witkowsk, B, Amaratunga C, Beghain J, Langlois A, Khim N, … & Menard D. (2014). A molecular marker of artemisinin-resistant Plasmodium falciparum Nature, 505(7481), 50-55. https://doi.org/10.1038/nature12876
- Maikentii JI, Umar RM, Ombugadu A, Aimankhu PO, Yusuf KS, Mamman AS, & Dakum YD. (2024). Malaria prevalence and associated risk factors among pregnant women attending Wadata and Lafia north primary health care centers Lafia, Nasarawa state, Nigeria. UMYU Scientifica, 3(3), 55-66. https://doi.org/10.56919/usci.2433.007
- Idris H, Anas U, Abdulmalik U, Jafar S. A., & Kabir, A. (2024). Prevalence of Plasmodium falciparum parasite among outpatients attending Muhammad Abdullahi wase teaching hospital in Kano state, Nigeria. Dutse Journal of Pure and Applied Sciences, 10(2a), 324-334. https://doi.org/10.4314/dujopas.v10i2a.31
- Lukuka, A., Mukomena Sompwe, E., Mukuku, O., Luboya Nku, M., & Mutombo, A. M. (2022). Rural malaria prevalence in the Democratic Republic of Congo. Tropical Medicine & International Health, 27(3), 310–321. https://doi.org/10.1111/tmi.13652
- Ouédraogo, A., Tiono, A. B., Diarra, A., Sanon, S., Yaro, J. B., Ouédraogo, E., … & Ouedraogo, A. (2020). Age-specific malaria prevalence in Burkina Faso. Malaria Journal, 19(1), 372. https://doi.org/10.1186/s12936-020-03446-5
- Mosha, D., Derua, Y. A., Simfukwe, E. T., Lyimo, I. K., Mwingira, V. S., Massaga, J. J., … & Mboera, L. E. G. (2021). Malaria transmission dynamics in Tanzania: Age-specific infection rates. Parasites & Vectors, 14(1), 289. https://doi.org/10.1186/s13071-021-04798-0
- Conrad, M. D., LeClair, N., Arinaitwe, E., Wamala, S., Kiggundu, M., Kamya, M. R., & Rosenthal, P. J. (2017). Comparative impacts over a decade of antimalarial drugs on the Plasmodium falciparum population in Uganda. Journal of Infectious Diseases, 216(5), 541-549. https://doi.org/10.1093/infdis/jix297
- Mensah, B. A., Aydemir, O., Myers-Hansen, J. L., Opoku, M., Hathaway, N. J., Marsh, P. W., Wilson, M. D., & Bailey, J. A. (2020). Genetic diversity and drug resistance markers in Plasmodium falciparum isolates from Ghana. Malaria Journal, 19(1), 276.
- Uwimana, A., Legrand, E., Stokes, B. H., Ndikumana, J. L. M., Warsame, M., Umulisa, N., Ngamije, D., Munyaneza, T., Mazarati, J. B., Munguti, K., Campagne, P., Kimenyi, J. C., Kabera, M., Bigira, V., Musanabaganwa, C., Rucogoza, A., Rusingiza, E. K., Halsey, E. S., Ramboarina, S., … & Menard, D. (2020). Emergence and spread of Kelch13 mutations in Rwanda. Nature Medicine, 26(10), 1602–1608. https://doi.org/10.1038/s41591-020-1005-2
- Amaratunga, C., Lim, P., Suon, S., Sreng, S., Mao, S., Sopha, C., Sam, B., Dek, D., Try, V., Amato, R., Blessborn, D., Song, L., Tullo, G. S., Fay, M. P., Anderson, J. M., Tarning, J., Fairhurst, R. M. (2016). Plasmodium falciparum founder populations in western Cambodia and their role in the emergence of artemisinin resistance. Nature Communications, 7, 119-130. https://doi.org/10.1038/ncomms11930
- Lu, F., Culleton, R., Zhang, M., Ramaprasad, A., Seidlein, L. von, Zhou, H., Zhu, Y., Tang, J., Liu, Y., Wang, W., Cao, Y., Xu, S., Gu, Y., Li, J., Kimani, J., Otieno, L., Oyibo, W., Phuc, B. Q., Toure, O. A., … & Yang, H. (2017). Emergence of Plasmodium falciparum kelch13 mutations in the Amazon Basin. Antimicrobial Agents and Chemotherapy, 61(3), e01905-16. https://doi.org/10.1128/AAC.01905-16
- Zubair AA, Gabriel AA, Ali AH., Karderam BD, Sagir AB, Simdung WK, Amina YB, Abubakar U, Muttaka A, & Muhammad RB. (2024). Molecular surveillance of artemisinin combination therapy resistance markers in Kelch 13 gene of Plasmodium falciparum isolates collected from Yobe State, Nigeria. Avicenna Journal of Clinical Microbiology and Infection. https://doi.org/10.34172/ajcmi.3524
- Kamau, E., Campino, S., Amenga-Etego, L., Drury, E., Ishengoma, D., Johnson, K., Mumba, D., Kekre, M., Yavo, W., Mead, D., Bouyou-Akotet, M., Apinjoh, T., Golassa, L., Randrianarivelojosia, M., Andagalu, B., Maiga-Ascofare, O., Amambua-Ngwa, A., Amaratunga, C., Amato, R., … & Kwiatkowski, D. P. (2015). K13-propeller polymorphisms in Plasmodium falciparum parasites from sub-Saharan Africa. The Journal of Infectious Diseases, 211(8), 1352–1355. https://doi.org/10.1093/infdis/jiu608
- Ndour, P. A., Gaye, A., Sy, M., Ndiaye, T., Siddle, K. J., Park, D. J., Deme, A. B., Mbaye, A., Dieye, B., Ndiaye, Y. D., Neafsey, D. E., Early, A., Farrell, T., Yade, M. S., Diallo, M. A., Diongue, K., Bei, A., Ndiaye, I. M., Volkman, S. K., & Ndiaye, D. (2020). Prevalence of kelch13 mutations in Senegal. Malaria Journal, 19(1), 301. https://doi.org/10.1186/s12936-020-03382-5
- Balikagala, B., Fukuda, N., Ikeda, M., Katuro, O. T., Tachibana, S.-I., Yamauchi, M., Opio, W., Uemura, H., Horii, T., Endo, H., Palacpac, N. M. Q., & Mita, T. (2021). Evidence of artemisinin-resistant malaria in Africa. New England Journal of Medicine, 385(13), 1163–1171. https://doi.org/10.1056/NEJMoa2101746
- Schmedes, S. E., Patel, D., Dhal, S., Kelley, J., Svigel, S. S., Dimbu, P. R., Adeothy, A.-L., Kahunu, G. M., Nkoli, P. M., Beavogui, A. H., Kariuki, S., Mathanga, D. P., Koita, O., Ishengoma, D., Mohamad, A., Hawela, M., Moriarty, L. F., Samuels, A. M., Gutman, J., Plucinski, M. M., Udhayakumar, V., Zhou, Z., Lucchi, N. W., Venkatesan, M., Halsey, E. S., & Talundzic, E. (2021). Plasmodium falciparum kelch13 mutations, 9 countries in Africa, 2014–2018. Emerging Infectious Diseases, 27(7), 1902–1908. https://doi.org/10.3201/eid2707.203230